Mutasjon og organer
Homeotiske mutasjoner og organidentitet
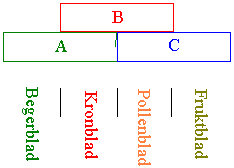
Blomsten hos angiospermene kan bestå av 4 konsentriske ringer med blader. Fra ytterst til innerst: grønne begerblad, fargete kronblad, hannlige pollenblad som lager pollen og hunnlige fruktblad som inneholder egg i en embryosekk. Johann Wolfgang von Goethe kom med hypotesen om at alle delene av blomsten var blad, beskrevet i boka Die Metamorphose der Pflanzen. Kronblad, begerblad, pollenblad, fruktblad kommer fra små grupper av celler i blomstringmeristemet, kalt primordier (l. primordium - begynnelse). Primordiene er i begynnelsen relativt udifferensierte, men på et seinere stadium får de organidentitet. Utrykket av blomstermorfologien (organidentiteten) er styrt av homeotiske gener. Loci på homeotiske gener bestemmer hvilken type blomsterblader som skal lages hvor. Homeotiske loci inneholder et DNA-bindende område kalt MADS-boks domene. MADS domene tilsvarer det man finner i Hox-gener i dyr (vertebrater), og som styrer dannelsen av lemmer og kroppsform. Navnet MADS har det fått fordi dette domene er blitt funnet i transkripsjonsfaktorer i gjær (mcm1), vårskrinneblom (ap3), løvemunn (defA) og hos menneske (srf). Homeotiske mutasjoner og homeotiske gener kan endre organformen slik at et organ dannes, men på feil sted eller til feil tid. Homeoboks gener koder for en klasse transkripsjonsfaktorer, kalt homeodomene proteiner, som regulerer gener som bestemmer skjebnen til celler og organers identitet. Doble og fylte blomster er laget og selektert av planteforedlere og gartnere. Blomster med mange farget kronblad er mer attraktive og populære enn dem med få. Det er homeotiske mutasjoner som forandrer pollenblad (pollenbærere) på villroser til kronblad i foredlete roser. Slike homeotiske mutasjoner hvor pollenblad er omdannet til kronblad kan man finne hos mange planter bl.a. hvitveis. De fleste studiene av blomsterutvikling er gjort med de to modellplantene vårskrinneblom (Arabidopsis thaliana) og løvemunn (Antirrhinum majus). Det er en hypotese om at 3 loci A, B og C bestemmer strukturen av blomster. A alene gir begerblad, A+B gir kronblad, B+C gir pollenblad og C gir fruktblad.

Eksempler på homeotiske mutasjoner. Øverst normal vill-type blomst. 1. Mutant hvor kronblad er omdannet til pollenblad og begerblad til fruktblad. 2. Mutant hvor pollenblad og fruktblad er omdannet til begerblad. 3. Mutant hvor kronblad er omdannet til begerblad og pollenblad til fruktblad.
Blomstringsmutanter kan oppstå eksperimentelt etter behandling med kjemiske mutagener eller radioaktiv stråling. Mutantene vises sjelden i M1-generasjonen (M-mutant). Derfor selvpollineres plantene og i neste M2-generasjon høstes interessante mutanter. Ca. 25% av avkommet etter selvpollinering vil være homozygote for en mutasjon. Ved gjentatte selvpollinering skaffer man seg stabile homozygote blomstringsmutanter. Hos Arabidopsis har man funnet genene APETALA1 (AP1) som tilsvarer A, APETALA3 (AP3) og PISTILLATA (PI) som tilsvarer B og AGAMOUS (AG) som tilsvarer C. Disse er overstyrt av et LEAFY gen. Mutasjon i LEAFY genet i Arabidopsis blokkerer for dannelsen av blomster. Det vegetative meristemet går ikke direkte over til blomster i vill-type Arabidopsis, men det transformeres først til et blomsterstandmeristem (inflorescens meristem) som lager en felles akse med brakter (tynt lite blad). I hjørnet (aksilen) av braktene lages det laterale meristemer som utvikles til blomstermeristemer. Tilsvarende blomstermutanter finner man hos Antirrhinum. Blomsten hos løvemunn (Antirrhinum) har bilateral symmetri. Cycloidea (cyc) og radialis (rad) er løvemunnmutanter som lager radiærsymmetriske blomster. Radiærsymmetriske blomster er evolusjonsmessig mer primitive og opprinnelige enn bilaterale og zygomorfe blomster. Dette gir mulighet til å studere hvordan evolusjonen har endret blomstersymmetrien. Hos revebjelle (Digitalis purpurea) kan man noen ganger observere at blomsterklasen blir determinert eller avsluttet med en stor radiærsymmetrisk blomst, kalt en pelorie.
Man har funnet homologe gener til LEAFY og MADS-boks gener i bregner og bartrær, og her har disse genene med dannelsen av reproduktive strukturer generelt og ikke blomster. Dette betyr sannsynligvis at disse genene var tilstede hos de første landplantene for 500 millioner år siden.
,