Sirkulær modell
Anta i et enkelt system at vi har en stabil bestand av predatoren rødrev og byttedyret hare. Ved tid=0 blir det satt ut et overskudd av hare, y=y0. Revene får mer mat og revebestanden øker, og disse dreper flere harer slik at harebestanden begynner å synke. Endring i revebestanden dx/dt og harebestanden dy/dt kan beskrives av differensialligningene:
\(\frac{dx}{dt}= y \)
\(\frac{dy}{dt}=-x\)
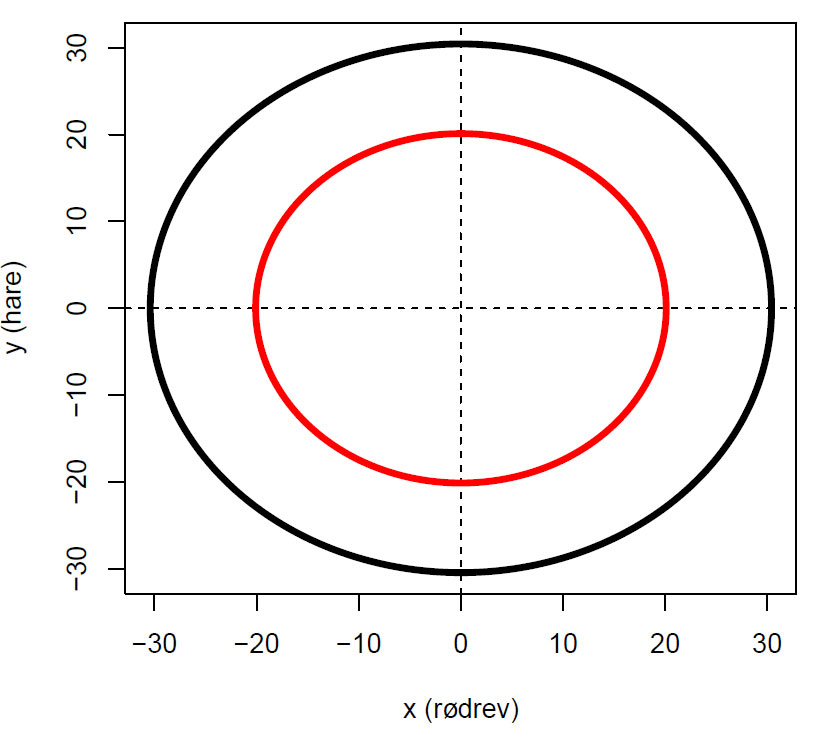
Fasediagram som viser lineær flyt omkring origo. Startbetingelser (x=5, y=30,svart) og (x=2, y=20,rød)
En lukket løkke i fasediagrammet indikerer periodiske svingninger, som vi kan observere i tidsseriediagrammet nedenfor. Hadde systemet vært stabilt hadde trajektoriet vist seg som bare ett punkt.
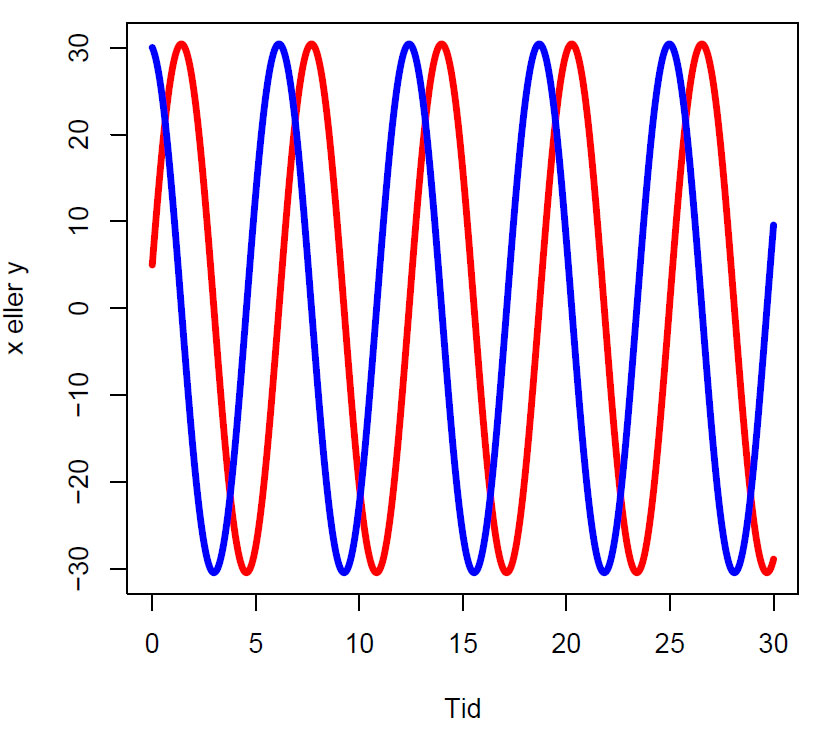
Tidsseriedigram for rødrev (x, rød) og hare (y, blå). Startbetingelser (x=5, y=30)Populasjonene gjennomgår sinusoscillasjoner med 90o ut av fase fra hverandre.
Lotka-Volterra modell
Lotka-Volterra ligningen er et par første ordens ikke-linære differensialligninger som beskriver dynamikken når to arter interagerer med hverandre: predator (P) og byttedyr (B).
Den italienske biologen Umberto D´Ancona hadde i 1926 tidsseriedata for fiskefangst i det nordlige Adriaterhavet med bl.a. prosenten av de forskjellige fiskeslagene på markedene i Trieste, Fiume og Venezia. Under første verdenskrig hvor fisket var redusert fant Ancona økt andel av predatorer (hai,skate,rokker). Ancona kontaktet Volterra som utviklet en modell for å kunne forklare fenomenet. Modellen er en forenkling av virkeligheten, men gir et innblikk i bruk av modeller. Det ble mulig å forklaring hvordan økt fiske kunne lede til mer fisk, ved at mengden predatorer også synker og det kompenserer for tapet av fisket (Volterras paradoks). Ved bruk av DDT mot skadeinsekter ble også insekter som predaterte på skadeinsektene også drept, slik at i noen tilfeller økte mengden skadeinsekter etter DDT-behandlingen !
Tankegangen ble presentert av både Alfred J. Lotka (1925) i studiet av kjemiske likevekter, og av Vito Volterra (1926). Endring i byttedyrbestand over tid er dB/dt og kan beskrives som:
\(\frac{dB}{dt}= rB\left(\frac{K-B}{K}\right)- bBP\)
Endring i bestanden av predatorer er dP/dt og kan beskrives som:
\(\frac{dP}{dt}= cBP-dP\)
Ligningene har en rekke konstanter:
r – vekstraten for byttedyr uten predator tilstede
b – effekten av predator på byttedyr
d – dødsrate for predator i fravær av byttedyr
c – effektivitet og formeringsrate hos predator
Predatorer dør i mangel på mat beskrevet av dP/dt=-dP.
I ligningen for byttedyr har vi satt inn et ledd (K-B)/K hvor vi antar at byttedyrbestanden ikke vokser uendelig, men når et metningsnivå med en bærekapasitet K.
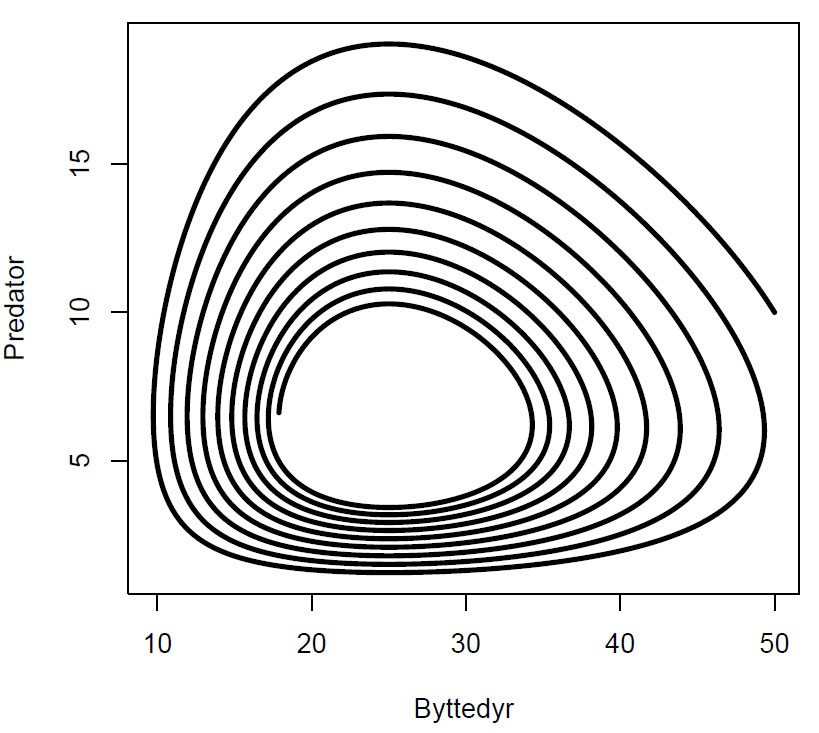
Fasediagram for r=0.2,K=500,b=0.03,c=0.02,d=0.5. Starter med 50 byttedyr og 10 predatorer Ved likevekt er(B,P)=(d/c,r/b)= (0.5/0.02, 0.2/0.03)=(25,7),som vi kan se av figuren er likevektspunktet med 25 byttedyr og 7 predatorer.
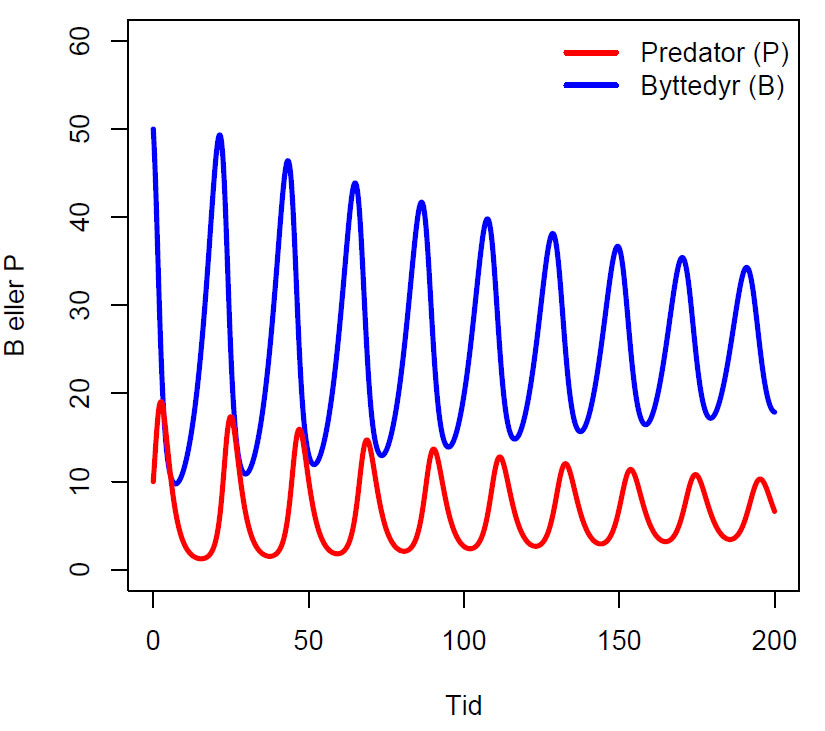
Lotka-Volterra ligningen for svingninger (oscillasjoner) i populasjoner av byttedyr og predator.
Konstanter: r=0.2,K=500,b=0.03,c=0.02,d=0.5. Start med 50 byttedyr og 10 predatorer. Etter en periode med svingninger i begge populasjonene vil de etter hvert stabilisere seg ved r/b= 0.2/0.03= ca 7 (predator) og d/c=0.5/0.02= 25 (byttedyr).
Hvis alle trajektoriene nærmer seg ett punkt er systemet stabilt. Kvasiperiodiske systemer har flere uavhengige periodiske spiraler rundt en torus.
Fasediagremmet har to akser, en for hver av de to populasjonene. Ethvert punkt i faserommet kan virke som initialbetingelser for systemet. I ethvert punkt på trajektoriet kan det trekkes en vektortangent som viser i hvilken retning systemet beveger seg og størrelsen på vektoren angir hvor stor forandringen er. Faserommet består av en mengde vektorer, danner et vektorfelt, som viser hvordan systemet oppfører seg. Trajektoriene beskriver hvilken vei et punkt beveger seg. Hvis vi starter å følge trajektoriet fra initialbetingelser (50,10) ser vi at predatorbestanden stiger, mens byttedyrbestanden synker. Dette skjer til vi når et maksimum og predatorbestanden begynner å synke. Vi finner nullkliner hvor dB/dt=dP/dt=0. I et faseportrett kan alle mulige trajektorier beskrives og kan betraktes som flytlinjene til en partikkel i en væske.
Hvis vi utelater leddet med bærekapasitet:
\(\frac{dB}{dt}= rB-bBP \)
\(\frac{dP}{dt}= cBP -dP\)
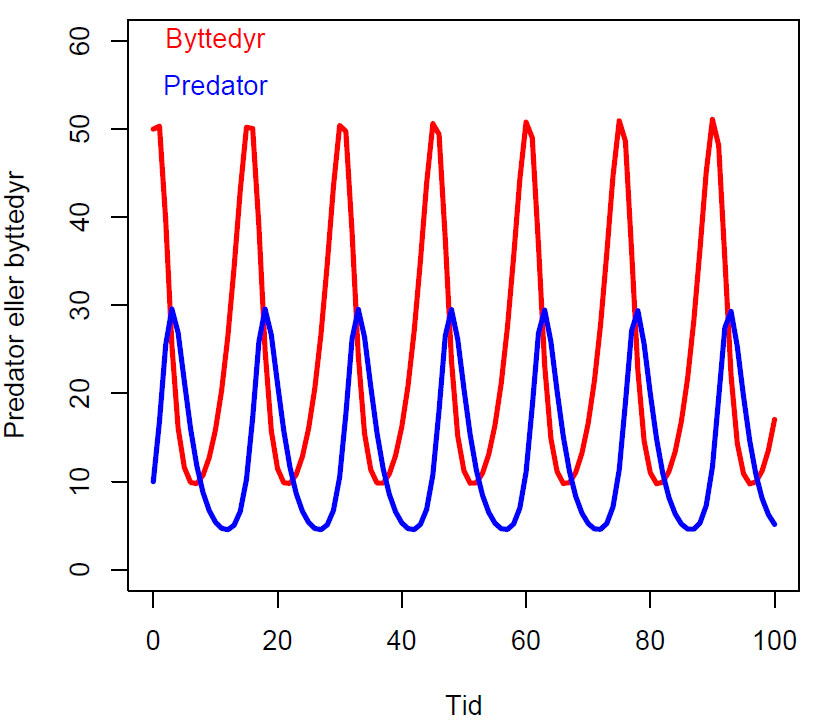
Tidsseriediagram Lotka-Volterra uten ledd for bærekapasitet, med parametere r=0.4,b=0.03,c=0.02,d=0.5, og initialbetingelser (50,10). Byttedyrbestanden svinger mellom 10 og 50,og predatorbestanden svinger mellom 5 og 30.
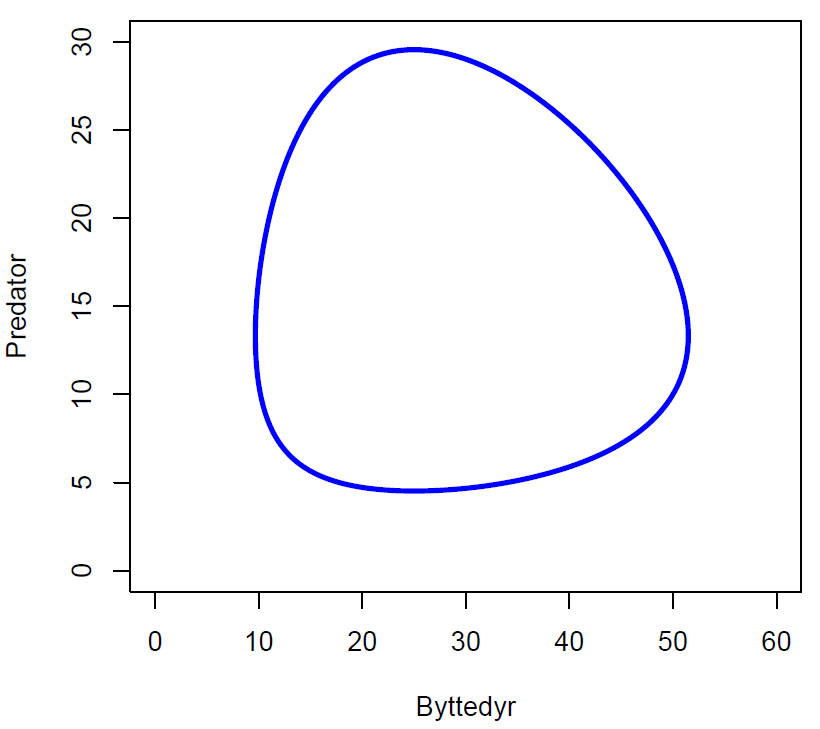
Fasediagram Lotka-Volterra uten ledd for bærekapasitet, med parametere r=0.4,b=0.03,c=0.02,d=0.5, og initialbetingelser (50,10).
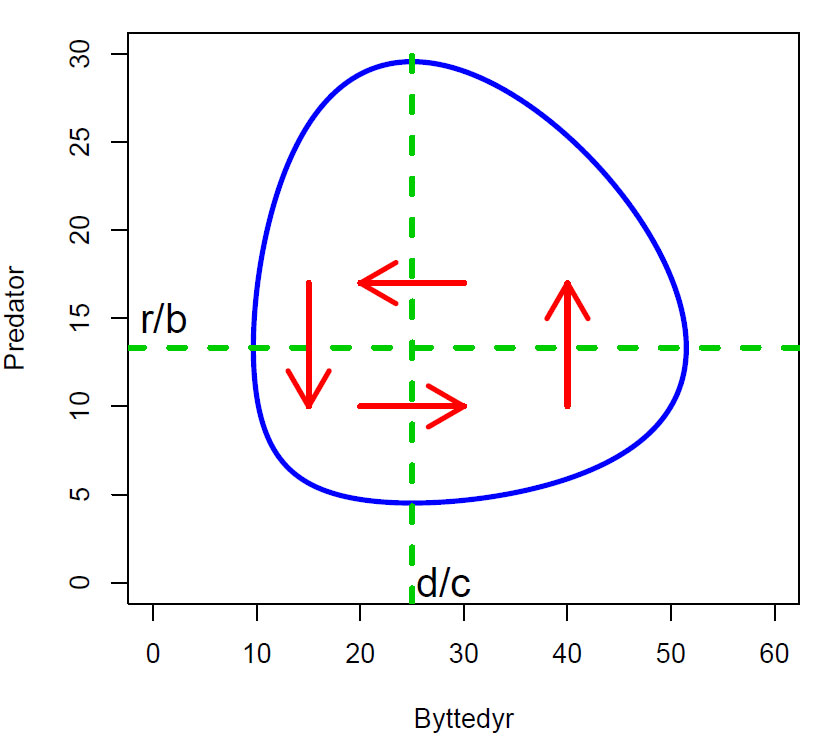
Lotka-Volterra fasediagram (faseportrett). I hvert punkt på trajektoriene (blå) kan det trekkes tangenter som viser i hvilken vei systemet beveger seg. Ved d/c og r/b er de deriverte lik 0 (d/c=25 byttedyr, og r/b=13.3) som beskriver gjennomsnittspopulasjonen av byttedyr og predatorer i en syklus. Deretter skifter systemet retning og beveger seg i retningen som de røde pilene viser, horisontale nullkliner for byttedyr og vertikale nullkliner for predatorer. Trajektoriene vender tilbake til utgangspunktet i en lukket geometrisk løkke.
Vi må først finne maksimums- og minimumspunktene til systemet hvor de deriverte er lik 0 og vi har likevektspunkter, nullvekstisokliner for predator og byttedyr:
\(\frac{dB}{dt}=\frac{dP}{dt}=0\)
Deretter undersøker man hvordan systemet oppfører seg rundt likevekstpunktene maksimum og minimum.
\(\frac{dB}{dt}= rB-bBP =B\left(r-bP\right)=0\)
\(\frac{dP}{dt}= cBP -dP=P\left(cB-d\right)=0\)
Som gir:
\(\left(B,P\right)= \left(\frac{d}{c}, \frac{r}{b}\right)\)
Jacobi-matrisen (J(B,P))med de partiellderiverte i predator-byttedyrligningen kan brukes til å undersøke stabiliteten ved et fast punkt. For ligningene:
\(\frac{dB}{dt}= rB-bBP \)
\(\frac{dP}{dt}= cBP -dP\)
Blir Jacobi-matrisen:
\(J\left(B,P\right)=\begin{bmatrix} r-bp &-bP\\ cB& cB-d \end{bmatrix}\)
Ved likevekt J(B,P)= J(0,0) blir denne:
\(J\left(0,0\right)=\begin{bmatrix} r & 0 \\ 0 & -d \end{bmatrix}\)
Vi finner egenverdiene lambda ved å løse:
\(\left| \begin {matrix} r-\lambda & 0\\ 0& -d-\lambda \end{matrix} \right|=0\)
Og finner egenverdiene
\(\lambda_ 1 = r\;\;\;\;\;\, \lambda_2 = -d\)
En positiv og en negativ egenverdi tilsvarer en ustabil sadelnode.
De to tilsvarende egenvektorene(ν) danner to akser:
\(J \cdot \nu= \lambda \cdot \nu\)
Generelt om stabilitet ved likevektspunkt og Jacobimatriser:
Hvis vi har et system med to differensialligninger med initialverdier (x0,y0) ved tid t=0:
\(\frac{dx}{dt}= f\left(x,y\right)\)
\(\frac{dy}{dt}= g\left(x,y\right)\)
Så kalles matrisen med de partiellderiverte for Jacobimatrise A.
\(A=\begin{pmatrix} \frac{\partial f}{\partial x} &\frac{\partial f}{\partial y} \\ \frac{\partial g}{\partial x} &\frac{\partial g}{\partial y} \end{pmatrix}\)
Jacobimatrise, egenverdier og egenvektorer benyttes i studiet av stabiliteten av likevektspunkter i differensialligningene.
Det kan være stabil eller ustabil likevekt. Systemet vokser eller synker i retning bestemt av egenvektorene og med hastighet bestemt av egenverdiene.
Lotka-Volterra konkurransemodell
Logistisk modell som gir mer intraspesifikk konkurranse ettersom populasjonsstørrelsen øker. Vi har to arter med endring i antall arter over tid (n1 og n2) med veksthastigheter r1 og r2 og bærekapasitet K1 og K2, hvor a12 og a21 er konkurransekoeffisienter, a12 er per capita effekten av art2 på art1. Dette er en symmetrisk modell:
\(\displaystyle \frac{dn_1}{dt}= r_1 \cdot n_ 1 \left(\frac{K_1-n_ 1+ a_{12 }\cdot n_2}{K_1}\right )\)
\(\displaystyle \frac{dn_2}{dt}= r_2 \cdot n_ 2 \left(\frac{K_2-n_ 2+ a_{21}\cdot n_1}{K_2}\right )\)
Hvis artene ikke interagerer dvs. a12 og a21 begge er lik 0 blir det en logistisk ligning. Hvis koeffisientene a12 og a21 begge er negative har vi mutualisme, hvis en er negativ og den andre er lik 0 er det kommensalisme. Hvis a12 og a21 begge er postive er det konkurranse mellom artene, og har de motsatt fortegn er det parasittisme.
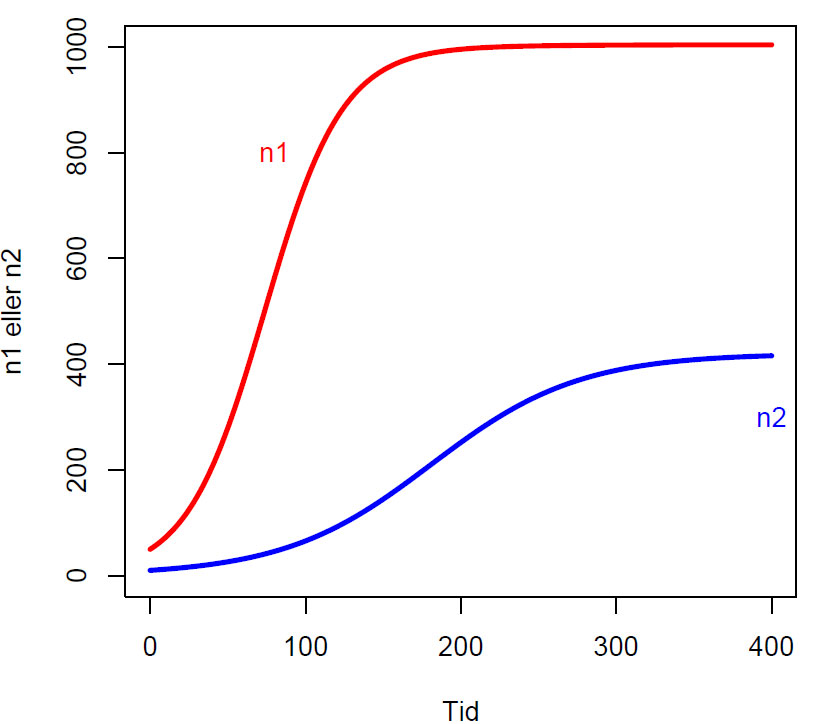
Figur Lotka-Volterra tetthetsregulert tidsseriemodell med parameterverdier r1=0.04, r2=0.02, K1=1000, K2=400, a12=0.01, a21=0.02)og initialverdier n1=50 individer og n2=10 individer.
Det finnes mange modeller for sammenhengen mellom endring i antall byttedyr over tid (dy1/dt) og antall predator over tid (dy2/dt) hvor K (bærekapasitet), a (dødsrate for predator i fravær av byttedyr, b, og c (effektivitet og formeringsrate hos predator) er konstanter:
\(\displaystyle \frac{dy_1}{dt}= y_1 \left ( 1-\frac{y_1}{K} \right ) - y_2 \cdot y_1 ^b\)
\(\displaystyle \frac{dy_2}{dt}= cy_2 \cdot y_1^b-ay_2\)
Rosenzweig-MacArthur predator byttedyrmodell
Rosenzweig-MacArthur predator byttedyrmodell hvor dy1/dt er endring bestanden av byttdyr og dy2/dt er endringen i predatorbestanden
\(\displaystyle \frac{dy_1}{dt}= r_1y_1 \left(1-\frac{y_1}{K_1}\right)-y_2 \frac{a_1y_1}{B + y_1}\)
\(\displaystyle \frac{dy_2}{dt}= y_2 \frac{a_2y_1}{B + y_1} - r_2 y_2\)
Med konstanter r1, r2, a1, a2, B og K1.
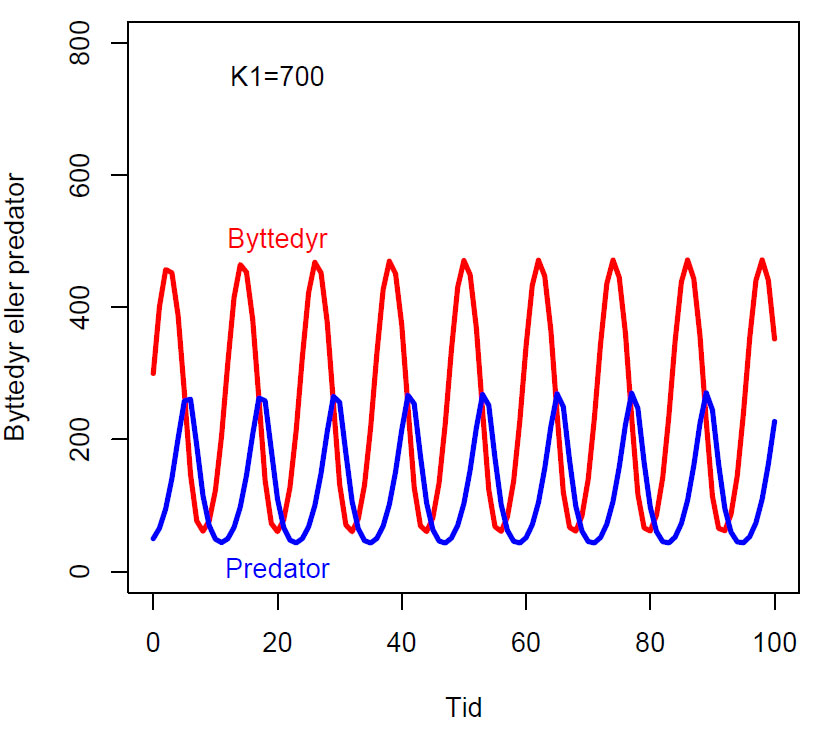
Fasediagram med parameterverdier a1=2, a2=2, r1=1, r2=1, K1=700, B=20
Endring i antall byttedyr over tid (dy1/dt) og antall predator over tid (dy2/dt) hvor K (bærekapasitet), a (dødsrate for predator i fravær av byttedyr, b, og c (effektivitet og formeringsrate hos predator) er konstanter:
\(\displaystyle \frac{dy_1}{dt}= y_1 \left(1-\frac{y_1}{K} \right) \)
\(\displaystyle \frac{dy_2}{dt}= cy_2y_1^b-ay_2 \)
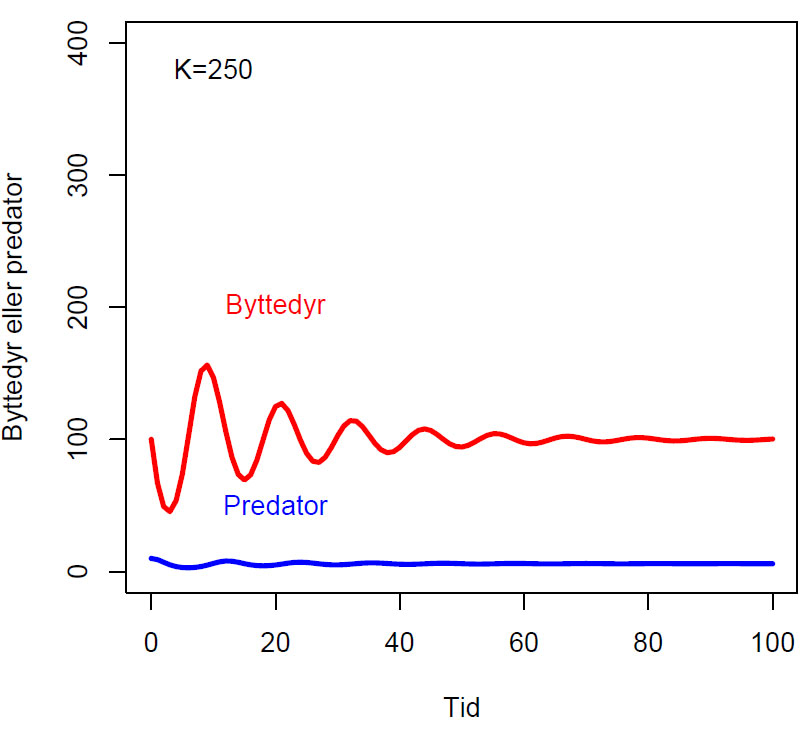
Fasediagram med parameterverdier b=0.5, K=250, c=0.1, a=1
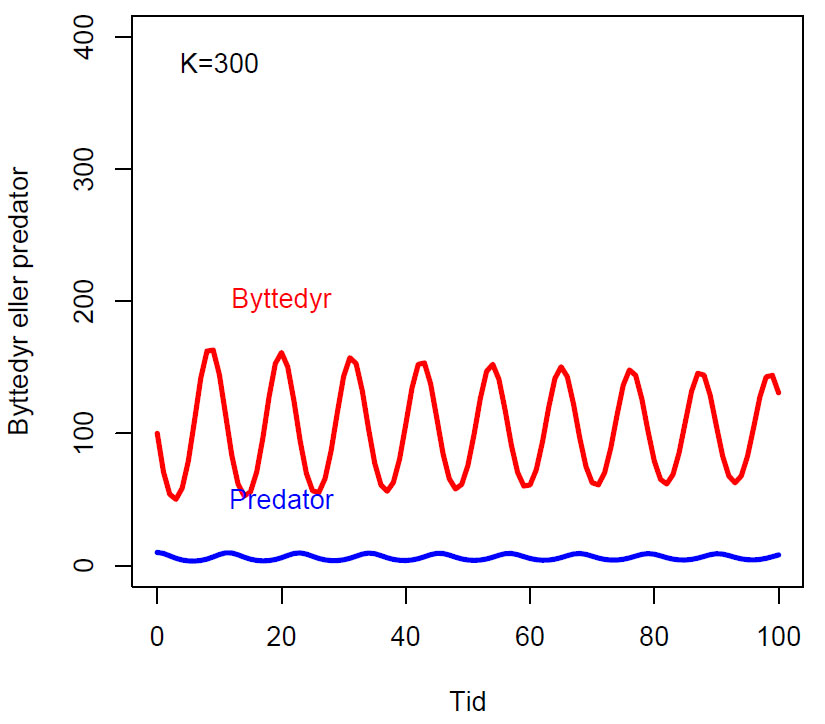
Fasediagram med parameterverdier b=0.5, K=300, c=0.1, a=1
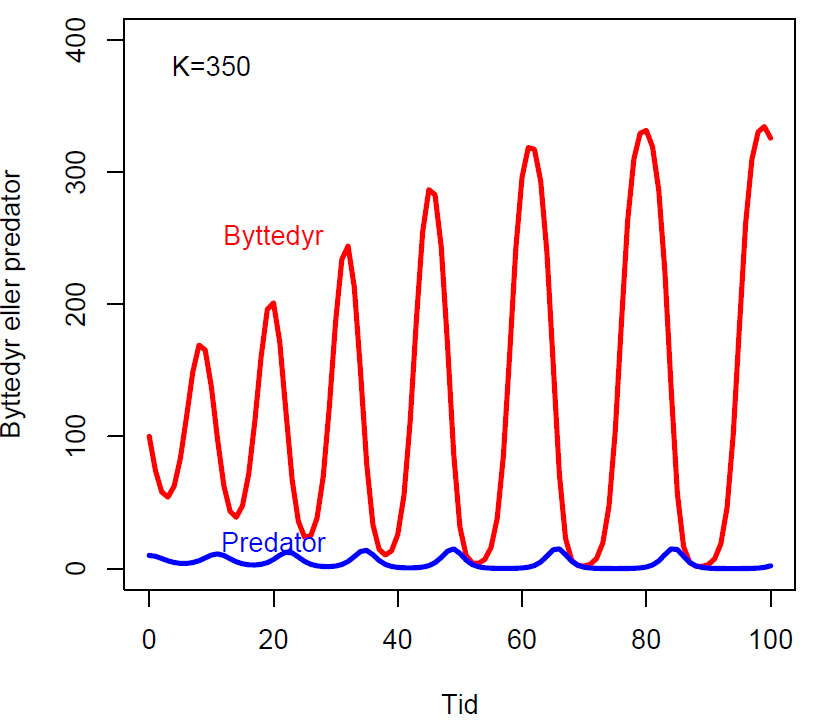
Fasediagram med parameterverdier b=0.5, K=300, c=0.1, a=1