Vannplanter
- Ferskvann
- Våtmarker
- Vannplantenes tilpasninger
- Tilpasning til hypoksis og anoksis
- Trykkdrevet gasstransport
- Løselighet av gasser i væske
Ferskvann
"Et svaberg skiller tang fra sivmorass og sjø fra myrvann, brunt som rustent jern." André Bjerke "Tjern ved kysten"
Oligotrofe sjøer er næringsfattige, klare og med liten produktivitet. Karakterarter er brasmegras (Isoetes), flotgras (Sparganium angustifolium) og botnegras (Lobelia dortmanna).
Eutrofe sjøer er næringsrike sjøer, og ligger ofte i områder hvor det er gjødselavrenning fra jordbruk. Karakteristiske arter er takrør (Phragmites australis), tjønnaksarter (Potamogeton), og langs land kan det vokse mannasøtgras (Glyceria fluitans) og strandrør (Phalaris arundinacea).
Dystrofe sjøer er brunfargede vann med mye oppløst humus (myrtjern), ofte med gul nøkkerose (Nuphar lutea) som en karakterart. Kalsium (Ca2+) kan danne kompleks med humusstoffer og gi utfellinger. Kalsiumfattig vann blir defor ofte brunfarget av humus.

Bilde. Gjengroing av tjern med flytetorv.
Dype sjøer har en lagdeling (stratifisering): De øvre vannmassene varmes opp av sola, utvider seg, og får mindre tetthet enn det kjøligere bunnvannet. Sommerprofilen har en varm epilimnion og en kald hypolimnion, atskilt av en termoklin (metalimnion) hvor det er stor temperaturforandring. Vind gir turbulens i det øvre vannlaget epilimnion. Det øvre laget tømmes for næringssalter om sommeren. Vår og høst er det mer lik temperatur i vannmassene og vannet blandes lettere av vind. Vinterstratifisering gir kaldt vann, og vannet like under isen har minst tetthet.
Det er transport nedover i vannet av nitrogen, fosfor og andre grunnstoffer i form av synkende celler, og avfall fra zooplankton som beiter på fytoplankton. Tilførsel av nitrogen og forsfor fra landbruk, kloakk eller ved næringsrikt vann fra hypolimnion kan gi algeoppblomstringer. Dette følges så av en oppblomstring av zooplankton som gresser på fytoplankton. I den trofogene sonen er det fotosyntese og syntese av organiske materialer og oksygen. I den trofolyttiske sonen skjer det nedbrytning og oksygen forbrukes. Det er en rekke fotosyntetiserende organismer i det akvatiske miljø: encellete og flercellete alger, makroalger og angiospermer.
Fytoplankton, produsentene i vannet, kan forekomme som encellede eller som kolonier. Blågrønnbakteriene kan danne klumper og kolonier (Anacystis), bunter med tråder (Aphanizomenon) eller lange kjeder (Anabaena). Algene kan inneholde gassvakuoler slik at de kan flytte seg i vannmassene. Diatoméene kan ikke bevege seg på denne måten. Planktongrønnalger forkommer som ubevegelige (Chlorella, Scenedesmus) eller med flageller (Chlamydomonas). I oligotrofe innsjøer er det lite fytoplankton f.eks. Staurastrum i svakt surt vann, og diatoméen Cyclotella i nøytralt og svakt basisk vann. I mesotroft vann og middels næringsrikt dominerer dinoflagellater av slekten Peridinium. I eutroft vann finnes Asterionella og Melosira om våren, og disse erstattes av Anacystis og Aphanizomenon om sommeren. En del av algefloraen består av celler og kolonier som lever på overflaten av steiner, bunnsedimenter og vannplanter. De festes av "slim" eller "limes" fast til substratet som en biofilm. Dette siste har stor betydning i elver og bekker.
Flyteplanter (lemnider) kalles planter som flyter fritt på overflaten av vannet f.eks. andmat (Lemna). Flytebladplanter (nymphaeider) kalles planter med rota i bunnen av vannet og assimilerende blad som flyter på vannoverflaten. Eksempler på slike er nøkkeroser, vanlig tjønnaks (Potamogéton natans), og piggknopp-arter (Spargánium). Planter med bladene nedsenket i vannet kalles vannskuddsplanter (eloeider). Slike er vasspest (Elodea) og tjønnaks-arter. Gras- og rusttjønnaks har både flyteblad og nedsenkete blad. Neddykkete planter med stive opprette skudd kalles kortskuddsplanter (isoetider, grunnskuddsplanter) og eksempler er botnegras (Lobelia dortmanna), brasmegras (Isoetes). Sumpplanter (helofytter, gr. helos - marsk; phyton - plante) kalles planter som vokser marsklandskap med røttene i vannmettet jord. Ålegrasartene (Zostera), havgras-arter (Ruppia) og dvergsivaks finnes i saltvann. Noen arter trives best i brakkvann (busttjønnaks, havgras, havfrugras). Tjønnaks (Potamogeton), vasspest (Elodea canadensis) og tusenblad (Myriophyllum) dominerer i alkalisk vann. Sure sjøer domineres av botnegras (Lobelia dortmanna), stivt brasmegras (Isoetes lacustris), og tjønngras (Litorella uniflora).
Fotosyntetiske bakterier har mindre betydning for primærproduksjonen i akvatiske systemer. Det er imidlertid 3 typer fotosyntetiske bakterier som lever anaerobt i vann, blant annet i vann nær kysten hvor bunnvannet danner et stabilt anaerobt saltvannslag dekket av ferskvann: grønne svovelbakteriene (slekten Chlorobium), purpur svovelbakterier (eks. slekten Chromatium) og purpur-ikke-svovelbakterier (eks. slekten Rhodopseudomonas). Makrofyttene er de makroskopiske plantene i vannet.
Refleksjon av lys fra en vannoverflate avhenger av solhøyde og bølger. Rødt lys absorberes raskest i rent vann og blått lys trenger dypest. Hvor mye lys som trenger ned i vannet avhenger av solhøyden, men også av turbiditeten i vannet. Lys brytes og spres av stoffer i vannet. Lys kan absorberes av gule pigmenter i vannet (gilvin), av fotosyntetiserende organismer og partikulært stoff (tripton). Langbølget varmestråling absorberes av de øverste mm av vannet. Forholdet mellom rødt/mørkerødt lys som påvirker fytokromsystemet øker lineært med dybden.
I vann er det en likevekt mellom karbonat, bikarbonat og karbondioksid avhengig av pH. I surt vann med pH lavere enn 5.5 vil mer enn 90 % av alt uorganisk karbon foreligge som karbondioksid (CO2). Ved pH mellom 7.5 og 9.5 er mer enn 90 % som bikarbonat (HCO3-). Totalinnholdet av uorganisk karbon blir lavt i surt vann siden alt foreligger som CO2. Akvatiske moser utnytter godt CO2 og dominerer i surt vann. I basisk vann kan det skje utfeller av kalsiumkarbonat (CaCO3).
Våtmarker
Våtmarker er områder hvor vannspeilet ligger over eller like over overflaten av jorda i større deler av året. Våtmarkene kan bli oversvømt og deles inn i:
1) Våtmarker hvor dekomponeringen går sakte og vi får en akkumulering av organisk stoff som i myr.
2) Våtmarker med liten akkumulering som gir sump. De vanligste artene slike steder er dunkjevle (Typha), takrør ( Phragmites), og Cyperus. Sumpplanter danner gjennomluftingsvev (aerenkym) og kan skille ut oksygen fra røttene som gir avsetninger av brunfargede Fe3+ oksider på røttene.
Vannplantenes tilpasninger
Vannplantene har følgende karakteristika:
1) Vannplantene har ofte heterofylli dvs. forskjellig bladform, avhengig av om bladene er over eller under vann. Bladene under vann er tynne og trådsmale. Dette gir store overflater og kortere diffusjonsvei for karbondioksid. I motsetning til blad over vann kan neddykkete blad være bare to cellelag tykke, og alle cellene i epidermis inneholder kloroplaster. Blad som flyter på overflaten har spalteåpningene mot oversiden.
2) Vann- og sumpplanter har et nettverk av luftkanaler (aerenkym) som danner et gjennomluftingsvev fra blad over vannet gjennom stengelen og ned til røttene. Aerenkymet sørger for oksygentilførsel til rot eller rotstokk. Oksygen som produseres under fotosyntesen i bladene samles i dette luftkanalsystemet og benyttes til respirasjon. Diffusjon og massestrøm er mekanismer som kan bidra til oksygentransporten internt i planten. Ofte har luftkanalsystemene ved nodiene en gjennomhullet plate med små porer. Luftvev kan dannes ved at intercellularrommene blir større eller ved at vev desintegrerer. I mangroveskog kan det dannes luftrøtter (pneumatoforer) som står opp over vannet.
3) Vannplantene har lite eller mangler fullstendig styrkevev. De mangler også lignin. Som regel er røttene lite utviklet. Høy tetthet i vann sammenlignet med luft reduserer behovet for styrkevev. Landplantene må investere store karbonressurser i styrkevev, sammenlignet med vannplanter.
4) En viktig faktor som begrenser fotosyntesen under vann er tilgangen på oppløst uorganisk karbon og lys. Vannplantene er tilpasset den lave diffusjonshastigheten for gasser i væskefase og de lave konsentrasjonene av karbondioksid (CO2) og oksygen (O2) i vann, sammenlignet med disse gassene i gassform. Avhengig av konsentrasjonene av CO2 og O2 vil forholdet mellom fotosyntese og fotorespirasjon variere. Dette gir stor variasjon i CO2- kompensasjonspunkt for en vannplante. Neddykkede vannplanter kan karakteriseres som skyggeplanter. Plantene bruker karbondioksid oppløst i vann som substrat for enzymet rubisko (ribulose 1,5-bisfosfat karboksylase), mens enzymet fosfoenolpyruvat karboksylase bruker bikarbonat som karbonkilde. Planter som vokser i surt vann som brasmegras og tjønngras har en form for C4-metabolisme med fosfoenolpyruvat karboksylase som CO2- fikserende enzym. Den lave Km-verdien fosfoenolpyruvat karboksylase har for bikarbonat gjør det letter å ta opp karon fra vannet, sammenlignet med ribulosebisfosfat karboksylase som bruker karbondioksid som substrat og har mye høyere Km-verdi. Ved høy pH i vannet vil planter som kan utnytte bikarbonat (HCO3-) ha en fordel. Disse vil også ha en lavere fotorespirasjon.
5) Ofte er det utfelling av ferrijern (Fe3+) på røttene som gjør dem brunfargede. Under anaerobe forhold i sedimentet med svart jord som inneholder FeS kan det bli så høye konsentrasjoner av ferrojern (Fe2+) at dette virker toksisk på planterøttene. Anaerobe forhold kan også gi toksiske konsentrasjoner av mangan.
6) Vannplanter kan overleve vinteren på bunnen av vann og innsjøer. Her kan de ha sine røtter, eller overvintre som vinterknopper (turioner). Daglengden via fytokromsystemet gir beskjed om når planten starter å lage turioner. Dannelse av turioner kan induseres av abscisinsyre.
Vannplantene har er rikelig tilgang på vann. De trenger derfor ikke spalteåpninger og beskyttelse mot vanntap i form av kutikula og tykkvegget epidermis. Blad som står i direkte i kontakt med vannmassene har en overflate dekket av karbohydratslim. Behovet for vanntransportsystem (xylem) hos vannplanter er betydelig minsket, og xylemtransporten fungerer bare i forbindelse med transport av næringssalter (grunnstoffer) som er tatt opp fra bunnsedimentene , men opptak av makro- og mikronæringsstoffer kan også skje via bladoverflatene. Høy varmekapasitet i vann gir mindre temperaturfluktuasjoner gjennom døgn og vekstsesong. Noen av vannplantene kan fotosyntetisere ned til 2 oC. Optimum er ca. 20 oC. Det kan ofte bli dårlig oksygentilgang for rotfaste vannplanter. Kraftig bevegelse i vannet kan skape et mekanisk stress. Mye tyder på at akvatiske angiospermer har vendt tilbake til vannet i sen evolusjonær tid. De er meget spesialiserte planter. Mange planter er "amfibier" som kan leve både i vann og på land. Planter med flyteblad finnes i alle sjøer uavhengig av om det er surt eller basisk, siden disse bladene står i direkte kontakt med luftas karbondioksid som landplantene. Karbonat kan ikke brukes i fotosyntesen. Det er bare enzymet fosfoenolpyruvat karboksylase som kan bruke bikarbonat (HCO3-) direkte, men spesielt ved høy pH blir HCO3- en viktig karbonkilde. Når HCO3- brukes blir det et overskudd på hydroksylioner (OH-). Opptak av karbonndioksid og bikarbonat øker derved pH slik at karbonat felles ut. Overgangen mellom karbondioksid og bikarbonat katalyseres av enzymet karbon anhydrase. pH kan stige meget høyt i de omgivende vannmasser ved høy fotosyntese. Det kan være slik at HCO3- tas opp på undersiden av bladet som derved blir sur, og hydroksylioner (OH- ) skilles ut på oversiden, som blir basisk. Spesielt sjøvann har høy konsentrasjon av HCO3- , og de marine algene bruker bikarbonat i sin fotosyntese. Neddykkede vannplanter har lav fotorespirasjon. De fleste kan utnytte bikarbonat som karbonkilde. Hos vasspest (Elodea canadensis) og Potamogeton arter vil det periplasmiske rom og grenselaget like ved bladoverflaten bli sur slik at likevekten mellom bikarbonat og karbondioksid forflyttes. Forsuringen skjer på den abaksiale siden av bladet og alkaliseringen på den adaksiale. Akvatiske C4 planter mangler kransanatomi, og C3 og C4 metabolismen er sannsynligvis lokalisert til forskjellige steder i cellen. All gassdiffusjon er i størrelsesorden 10.000 ganger mindre i vann enn i luft, og tykkelsen på grenselaget omkring neddykkete planter varierer fra 100 til 500 Botnegras (Lobelia dortmanna), tjønngras (Litorella uniflora) og brasmegras (Isoetes sp.) benytter seg av at det ofte er høyere konsentrasjon av karbondioksid i bunnsedimentene enn i vannet. Disse plantene vokser som rosetter og har stor rotutvikling og velutviklet luftkanalsystem fra røtter til skudd. Siden transporten av CO2 fra bunnsedimentene skjer ved diffusjon kan ikke bladene være for lange. Noen akvatiske makrofytter kan også ha blad over vann i visse stadier av livssyklus. Dette gir økt CO2-tilgang, og økt transpirasjon fra disse bladene vil også gi økt næringsopptak fra bunnsedimenter. Vannplantene har spesialisert seg til en visse vanndybde, selv om en plante som nøkkerose er svært fleksibel når det gjelder vannstand. Dypere enn 1 m blir det et meget stort forhold mellom rødt og mørkerødt lys. Fytokromsystemet må respondere annerledes hos planter som lever helt neddykket. For vannplanter med flyteblad blir fytokromlikevekten viktig. Ved pH rundt 7 finner vi blandete populasjoner. Utfellinger av kalsiumkarbonat (CaCO3) på vannplanter skyldes en interaksjon mellom planten og fysiokjemiske egenskaper til vannet rundet planten. Hos tjønnaks tas bikarbonat opp på undersiden og hydroksylioner (OH-) skilles ut på oversiden. Nitrogen, fosfor og silisium har stor betydning for produktiviteten i akvatiske systemer. mm. For landplanter er dette laget bare noen få mm.CO2 konsentrasjonen i vann overstiger sjelden 350 mM. Konsentrasjonen av bikarbonat varierer fra 0 til 5 mM i hardt vann. Fotosyntesen er halvmettet ved CO2 konsentrasjon 60-300 mM og mettet ved 350-650 mM. For bikarbonatbrukere er den halvmettet ved 0.5-0.8 mM og mettet ved 1.5-2 mM.
Tilpasning til hypoksis og anoksis
Selv om eukaryotene trenger oksygen behøver det ikke bety at de trenger konstant tilførsel av oksygen. Oksygenkonsentrasjonen som deler den aerobe fra den anaerobe verden kalles Pasteurpunkt. Vannplanter med røtter i bunnsedimenter kan bli utsatt for lave oksygenkonsentrasjoner (hypoksis) og perioder uten oksygen (anoksis), selv om de har aerenkym. Glykolysen skjer uten oksygen, men gir lavt utbytte med ATP, sammenlignet med trikarboksylsyresyklus og elektrontransportkjeden. Ved dårlig tilgang på oksygen til røttene (hypoksis) kan trikarboksylsyresyklus bli hemmet. Under anaerobe forhold (anoksis) skiftes karbohydratmetabolismen i rotcellene over til fermentering. Ved fermentering (etanolgjæring) kan det lages etanol katalysert av enzymene pyruvat dekarboksylase og alkohol dehydrogenase. Ved melkesyregjæring vil melkesyre lages av laktat dehydrogenase. Melkesyre (laktat) vil gi forsuring av cellene og kan gi skade ved acidose hvis det ikke skjer buffring. Etanol er delvis fettløselig og kan diffundere gjennom membraner, og plantene kan på denne måten unngå skade. Vannplantene har evne til bedre å tolerere hypoksis og anoksis enn landplantene. Anoksis og hypoksis vil gi lavere innhold av ATP, og mer ADP, AMP og adenosin i cellene, og dette kan virke som et signal til å øke karbongjennomstrømningen i glykolysen. Planter som tåler godt dårlig oksygentilgang er havsivaks (Scirpus maritimus), ris (Oryza sativa), takrør (Phragmites communis), og bred dunkjevle (Typha latifolia).
Trykkdrevet gasstransport
Neddykkete fastsittende vannplanter frakter oksygen til røttene via massestrøm. Dette gjøres ved å opprettholde en trykkforskjell i lakunesystemet (aerenkym) med en forskjell i atmosfæretrykk i blad og rot. Denne forskjellen i trykk oppnås ved transpirasjon og lufttrykk, og krever nærvær av små porer i aerenkymet med radius mindre enn 0.1 mm. Disse porene hindrer massestrøm og gir økning i trykk. I de yngste bladene blir lufttrykket høyere enn på utsiden, og oksygenrik luft følger en trykkgradient ned til rot eller rotstokk. Eldre blader har porer med større diameter som ikke skaper så stort luftovertrykk. Metanrik luft fra metanogene bakterier i bunnsedimentene strømmer ut av de eldre bladene og ut i lufta.. Oksygen har relativt liten løselighet i vann og når oksygenkonsentrasjonen øker ved fotosyntesen øker partialtrykket i de unge bladene som kommer i tillegg til økning i vanndamptrykket. Nøkkerose har en trykkdrevet ventillasjonsmekanisme for å frakte oksygen ned til røttene. Oksygen beveger seg fra unge blad gjennom rhizomene og ut gjennom de eldste bladene. Vanndamptrykket i bladet er større enn i omgivelsene. Temperaturøkning i bladene om dagen gir vanndamp som fortynner andre gasser og lager en diffusjonsgradient. Metan som lages av anaerobe bakterier i sedimentet på bunnen av vannet finnes i aerenkymet i hele planten om natten. Om dagen finnes bare metan i stengelen på gamle blad
Løselighet av gasser i væske
Løseligheten av karbondioksid (CO2) og dioksygen (O2) avhenger av temperatur, partialtrykk av gassen, type gass og løsningsmiddelet. For tynne løsninger sier Henrys lov at ved konstant temperatur vil mengden av gass som løses i et gitt volum væske være proporsjonal med partialtrykket til gassen. Gasser er generelt mindre løselige ettersom temperaturen øker. Løseligheten av oksygen avhenger ikke av pH slik som tilfellet er for karbondioksid. Siden pH stiger i stroma under fotosyntesen, og derved forflytter likevekten mot bikarbonat, må zinkenzymet karbon anhydrase sørge for et høyt partialtrykk av CO2 der hvor fotosyntesen skjer.
Vannplanter har behov for mekanismer som kan oppkonsentrere karbondioksid og det finnes pumper som utveksler bikarbonat og CO2 mot protoner (H+) og natrium (Na+).
Løselighet av gasser i væske er avhengig av fire faktorer:
- 1) Trykket til gassen og løseligheten øker med økende trykk.
- 2) Temperaturen på løsningsmiddelet. Løseligheten minsker med økende temperatur.
- 3) Hvor mye salt det er løst i væsken. Løseligheten minsker med økende saltinnhold.
- 4) Absorbsjonskoeffisienten til gassen.
Henrys lov kan også uttrykkes som at partialtrykket til en gass over løsningen er proporsjonal med molbrøken til gassen i løsningen. Konsentrasjonen av CO2 og O2 blir mindre i sjøvann enn i ferskvann ved samme trykk og temperatur. CO2 og O2 har også forskjellig løselighet ved forskjellig temperatur, og løseligheten av O2 er større i forhold til CO2 ved økende temperatur. Dette siste er en medvirkende årsak til at fotorespirasjonen øker med økende temperatur idet tilgangen til substratet oksygen for rubisko øker i forhold til substratet CO2.
Likevekten mellom karbondioksid og bikarbonat er gitt ved:
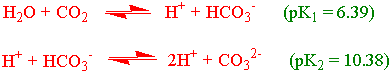
pK1 verdien for bikarbonat endrer seg med temperaturen slik at den er 6.392 ved 20 oC og 6.312 ved 40 oC dvs. ca. endring på 0.004 per oC.
Bruker vi Henderson-Hasselbalch ligningen får vi:
![]()
De finnes mer karbondioksid i vann enn det man forventer ut fra Henrys lov siden karbondioksid foreligger i andre former som også virker som et buffersystem i vann. Det er en kalsium-karbonsyrelikevekt: I vann med mye Ca2+ kan kalsiumkarbonat felles ut på bladene når plantene tar opp karbondioksid.