Fotosyntese
Reparasjon av skader i fotosynteseapparatet
Fotosynteseapparatet i kloroplastene kan skades på forskjellige måter:
- 1) For mye lysenergi inn i fotosystene i forhold til behovet.
- 2) Fotodynamisk produksjon av singlett oksygen og oksygenradikaler.
Plantene har beskyttelsessystemer som har til oppgave å minske lysabsorbsjonen og fjerne overskuddsenergi.
Ved lave lysflukser er det stor sannsynlighet for at de fleste tilgjengelig lyskvantene kan bruker til å eksitere klorofyllmolekyler, men ved høy lysfluks kan ikke energien til alle tilgjengelige lyskvantene benyttes til å lagre kjemiske energi. Planter som blir utsatt for mye solenergi kan redusere energimottaket ved å endre bladvinkel i forhold til lyset. Det kan også ha lysreflekterende overflate. Overskuddsenergi kan fjernes som fluorescens eller varme. VAZ-syklus som består av en syklus av karotenoidene violoxanthin, antherxanthin og zeaxanthin fjerner overskuddsenergi fra fotosyntesen og fra singlett oksygen som varme.
Det primære skadestedet ved fotoinhibering er D1-proteinet. For å hindre skade blir dette proteinet bygget raskt ned av protease og lages deretter på nytt slik at proteinet er i meget rask omsetning.
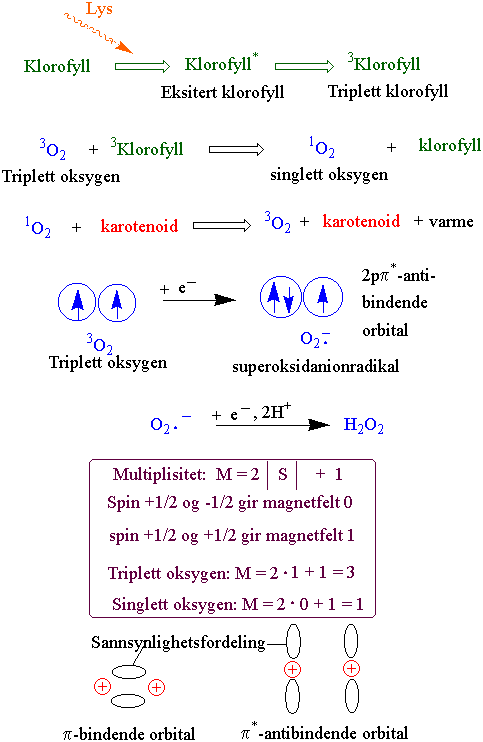
Fotodynamisk aktivering av oksygen
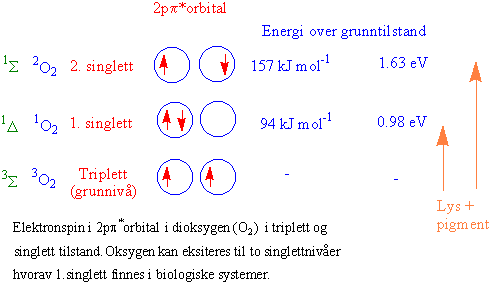
Fotodynamisk aktivering av oksygen kan skje ved at noen av klorofyllmolekylene omdannes til et triplettstadium som har lang nok levetid til å få overført energi til oksygen som derved omdannes til singlett oksygen som har relativt lang levetid i den hydrofobe membranen. I tillegg til at klorofyllmolekylet selv kan bli fotooksidert kan singlett oksygen oksidere umettede fettsyrer ved å danne peroksider. Karotenoider kan fjene overskuddsenergien i form av varme. I tillegg skjer beskyttelse med a-tokoferol og askorbinsyre. I C3 planter kan fotorespirasjon være en viktig beskyttelsesmekanisme ved å opprettholde elektrontransporten i karbonkjeden ved å slippe ut CO2. Katalase finnes ikke i kloroplasten. O2- avgiftes med Cu-Zn superoksid dismutase som gir hydrogenperoksid H2O2 som reduseres til vann via askorbat peroksidase-glutathion reduktase syklus.
{kind=link}
{kind=link}
{kind=link}
VAZ-syklus (xanthofyllsyklus) fjerner overskuddsenergi som varme
Karotenoider som ligger i thylakoidmembranene kan både virke som lyshøstende pigment, men har også en funksjon i fotobeskyttelse av fotosynteseapparatet. Karotenoidene virker som en sikkerhetsventil som fjerner overskudd av eksitasjonsenergi som kommer inn i fotosyntesene ved høy lysfluks. Karotenoidene virker i denne fotobeskyttelsen ved å fjerne eksitasjonsenergi fra klorofyll som varme.
Xanthofyllsyklus er en syklus som gir en lysavhengig og reversibel minskning av violaxanthin i grønne blad. Violaxanthin omdannes til zeaxanthin via intermediatet antheraxanthin. VAZ-syklus fjerner overskudd av eksitasjonsenergi som varme og beskytter planten mot fotooksidative skader under lysstress. Enzymet som katalyserer overføringen av violaxanthin til zeaxanthin er en del av thylakoidmembranen inn mot lumen og har et pH optimum ved pH 5.2. Under elektrontransporten i fotosyntesen pumpes H+ inn i lumen. Hvis syntesen av ATP går saktere så øker konsentrasjonen av H+ inne i lumen og mengden zeaxanthin øker. Enzymet de-epoksidase overfører violaxanthin til zeaxanthin med askorbat som kofaktor. Hvis derimot ATP syntesen er høy blir det mindre pH gradient og reaksjonen skjer fra zeaxanthin til violaxanthin katalysert av en epoksidase som bruker NADPH.
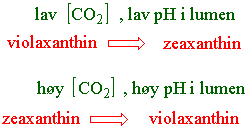
Det er indikasjoner på at det kan være en sammenheng mellom konsentrasjonen av zeaxanthin og åpning og lukkeing av spalteåpninger. Kloroplasten i mørkespirte koleoptiler inneholder violaxanthin og antheraxanthin, mens i lys dannes zeaxanthin. Dette kan muligens skyldes at kloroplastene i koleoptilen har en spesiell funksjon.