Mineralnæring
Fosfor
Fosfat behøver ikke reduseres før assimilering inn i organiske forbindelser, i motsetning til nitrat og sulfat. Etter å ha blitt tatt opp i plantene som H2PO4- forblir fosfat som uorganisk fosfat (Pi, i="inorganic") eller fosfat kan bli forestret til en hydroksylgruppe (CO-P) som en fosfatester eller energirik polyfosfat f.eks. pyrofosfat (PPi). I mange reaksjoner er fosfat et substrat eller produkt. Vakuolen er lagringssted for ikkemetabolsk fosfat. Fosfor inngår en rekke steder i metabolismen.
Fosfat i nukleinsyrene DNA og RNA
Det er fosfor som gir de sure egenskapene til nukleinsyrene. Fosfatgruppen vil ha negativ ladning ved nøytral pH. Et nukleotid består av en base, sukker og fosfat, og fosfat er bundet til sukker i en esterbinding.
Fosfatestere og energimetabolisme
Fosfat inngår i energimetabolismen. Hydrolyse av esterbindingen til fosfat frigir store mengder energi fra høyenergimolekyler. Hydrolyse av ATP katalysert av enzymet ATPase gir ca. 30 kJ mol- 1. En fosforylgruppe i ATP kan overføres til andre molekyler. Andre energirike fosfatestere er fosfoenolpyruvat (-62 kJ mol-1), pyrofosfat (-28 kJ mol-1) og glukose-6-fosfat (-14 kJ mol-1). ATP lages ved substratfosforylering i respirasjonen eller ved fotofosforylering i fotosyntesen katalysert av enzymet ATPsyntase.
Fosfolipider i biomembraner
Fosfodiestere i form av fosfolipider er vanlig i alle biomembraner f.eks. som fosfatidylcholin (lecithin). Fosfolipidene har en hydrofob hale dannet av to fettsyrer som vender inn i membranen og et hydrofilt hode som vender ut mot cytosol eller stroma.
Fosfor og signaloverføring
Fosfor inngår i signaloverføringsveier i metabolismen ved fosforylering og defosforylering av proteiner katalysert av henholdsvis protein kinaser og defosforylaser. GTP og GDP inngår i G- proteiner.
Transporten av triosefosfater ut av kloroplasten skjer via en fosfattranslokator i bytte mot fosfat. Fosfattranslokatoren sitter i den indre kloroplastmembranen. Syntesen av stivelse hemmes av høye konsentrasjoner av fosfat. Enzymet for syntese av stivelse i kloroplasten Enzymet ADP-glukose pyrofosforylase som lager stivelse i kloroplastene hemmes av uorganisk fosfat og aktiveres av triosefosfater. Stivelse fosforylase er et reversibelt enzym i plastider:
![]()
Fytat - et lager med fosfat og andre grunnstoffer
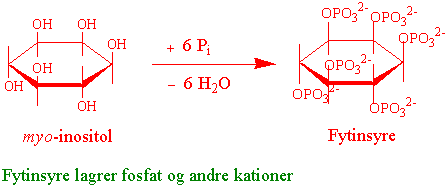
Fosfat lagres som fytat (fytin) i pollen, frø og korn. Fytat er et kalsium-magnesiumsalt av fytinsyre. Fytinsyre lages fra myo-inositol ved at hydroksylgruppene er forestret med fosfat. Fytinsyre (inositol heksafosforsyre) har også høy affinitet til andre grunnstoffer som jern og zink. Fytat spaltes av enzymet fytase. Syntesen av fytat i frø regulerer også stivelsesyntese ved å senke fosfatkonsentrasjonen via reaksjonen med fytat.
Fosfor i jord og mangelsymptomer
Fosfor er sterkt bundet i jorda, til jern og aluminium i sur jord og til kalsium i mer basisk jord. I moldjord finnes en stor del av fosforreservene i organisk form. Ved dårlig tilgang på fosfat kan planterøttene skille ut enzymet fosfatase til jorda hvor enzymet hydrolyserer organiske fosforforbindelser. Mykorrhiza kan lette opptaket av fosfor i røttene. Ved mangel på fosfor hemmes veksten av skuddet, men rotveksten hemmes ikke i samme grad. Fosfat kan reallokeres fra de eldste til de yngste delene av planten via floemet slik at fosformangelsymptomer viser seg først på de eldste bladene. Planter med fosformangel kan bli rødfarget av anthocyanin analogt til nitrogenmangel. De yngste delene av planten har dypgrønn klorofyllfarge, og skiller seg således fra nitrogenmangel hvor plantene blir lysegrønne og klorotiske.