Mineralnæring
Nitrogen
- Nitrogenassimilasjon
- Ammoniumassimilasjon
- Biologisk nitrogenfiksering
- Nitrogenfiksering hos planter og rhizobia
Nitrogenassimilasjon
Nitrogen er det grunnstoffet fra jorda som i størst grad regulerer veksten av planter. Nitrat og ammonium er hovedkildene til uorganisk nitrogen som tas opp av planterøttene. Nitratassimilasjonen skjer i både røtter og skudd. Noen planter i symbiose med mikroorganismer kan ta opp dinitrogen fra lufta. Aminosyrer i jorda og urea er andre nitrogenkilder plantene kan utnytte.
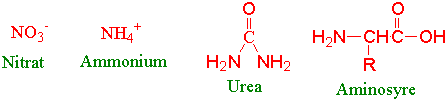
Nitrat reduseres til ammonium ved hjelp av to enzymer:
- 1) Nitrat reduktase, en to-elektron reduksjon av nitrat til nitritt.
- 2) Nitritt reduktase, en seks-elektron reduksjon av nitritt til ammonium.
Det er nær kobling mellom nitrogenmetabolisme og fotosyntese, siden fotosyntesen bidrar med reduksjonskraft (redusert ferredoksin eller NAD(P)H) som trengs for å redusere nitrat til ammonium. Nitrat reduktase i cytoplasma inneholder de prostetiske gruppene flavin adenin dinukleotid (FAD), cytokrom b557 og molybden. FADH2 i nitrat reduktase kan også redusere andre elektronekseptorer som Fe3+-citrat. Det er sjelden nitritt akkumuleres i planten under normale betingelser noe som tyder på synkronisering av de to enzymaktivitetene.
![]()
Nitritt som er produktet fra nitrat reduktase går inn i plastider og omdannes til ammonium av nitritt reduktase. Nitritt reduktase inneholder sirohem og finnes i kloroplaster i bladene eller i plastider i røttene.
![]()
C4-planter har både nitrat reduktase og nitritt reduktase i mesofyllcellene, men de finnes ikke i slirecellene. ofte en sammenheng mellom aktiviteten til nitrat reduktase og mengden redusert nitrogen i blad. Nitritt reduktase inneholder jernporfyrin. Genet for nitrat reduktase reguleres av nitratkonsentrasjon, lys og karbohydrater. Nitrat reduktase kan fosforyleres og defosforyleres analogt med enzymet sukrosefosfat syntase. I mørke aktiveres en serin protein kinase og nitrat reduktase inaktiveres. Planter som ikke har hatt tilgang til nitrat har et lavkapasitets konsitutitvt opptakssystem for nitrat, men ved nitrattilførsel vil i løpet av 20 minutter et høykapasitetssystem etableres. Klorat som brukes som ugrasdreper virker som en analog til nitrat.
Nitrat kan lagres i vakuolen hos nitrofile arter som meldestokk (Chenopodium album) og brennesle (Urtica dioica). Planter som vokser ved lave lysflukser i drivhus om vinteren kan inneholde høye konsentrasjoner med nitrat. Spinat inneholder mye nitrat og spinat bruker nitrat som osmoregulator hvor konsentrasjonen kan komme opp i 100 mM.
Ammoniumassimilasjon
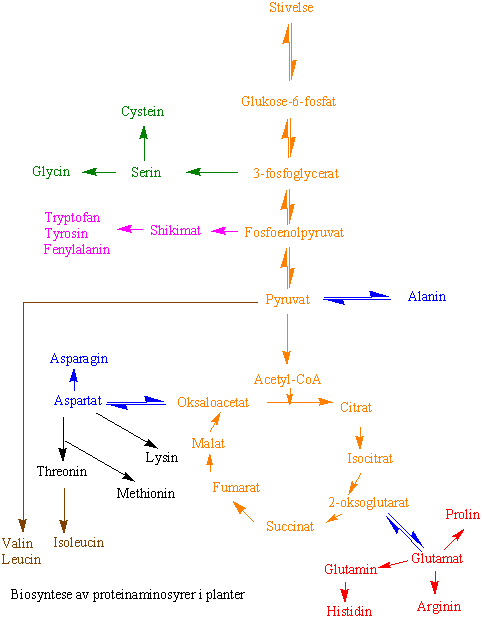
Uorganisk nitrogen i form av ammonium assimileres via enzymene glutamin syntetase (GS) og glutamin-oksoglutarat aminotransferase (GOGAT) inn i organiske forbindelser i plantene som aminosyrene glutamin og glutamat. Nitrogen kan deretter overføres til andre aminosyrer og brukes til syntese av nukleinsyrer, klorofyll og andre forbindelser som inneholder nitrogen.
Glutamin syntetase katalyserer omdanning av glutamat til glutamin i en ATP-avhengig reaksjon. Glutamin syntetase som har høy affinitet for ammonium (lav Km-verdi):

I blad er det to isoenzymer av GS, en i cytosol og en i kloroplasten. GS finnes også i ledningsvevet. GS aktiveres av høy pH og høye konsentrasjoner av ATP og Mg2+.
Det neste trinnet i ammonium assimilasjonen, glutamat syntase (GOGAT), katalyserer overføringen av amidgruppen (-NH2) fra glutamin til 2-oxoglutarat - et produkt fra Krebs-syklus. Glutamat syntase også kalt glutamin:2-oxoglutarat aminotransferase (GOGAT) lager glutamat med hjelp av redusert ferredoksin i blad:
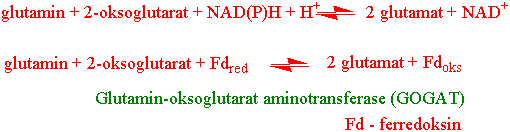
NADH-GOGAT er vanligst i ikke-fotosyntetiserende vev i røtter og ledningsstrenger. GS/GOGAT tar også hånd om de store nitrogenmengdene som frigis under fotorespirasjon. Plantene innholder to typer GOGAT, et NADH-avhengig (NADH-GOGAT) og et ferredoksinavhengig enzym (Fd- GOGAT).
Ammonium som er blitt assimilert kan overføres til ketosyrer via transaminering, for deretter å bli overført til andre proteinaminosyrer. Det finnes flere forskjellige aminotransferaser, også kalt transaminaser, f.eks. aspartat aminotransferase, alanin aminotransferase, greinete aminosyrer aminotransferase o.s.v. Transaminaser trenger pyridoksalfosfat (vitamin b6) som kofaktor. Enzymene finnes i cytoplasma og alle organellene i cellen. I proteinsyntesen blir de forskjellige aminosyrene koblet sammen med peptidbindinger (R1-CO-NH-R2). Proteiner er polypeptider som består av mer enn 100 aminosyrer.
{kind=link}
I mitokondrier og kloroplaster finnes enzymet glutamat dehydrogenase, som man opprinnelig trodde hadde ansvaret for assimilasjonen av ammonium. Det er imidlertid mer rimelig at enzymet har med nedbrytning av glutamat

Biologisk nitrogenfiksering
Mikroorganismer bruker tre livsstrategier for for å fiksere dinitrogen fra lufta: symbiotisk, assosiativ og frittlevende. Biologisk nitrogenfiksering er energikrevende og trenger minst 960 kJ mol- N som fikseres. Nitrogenase er et enzym som bare finnes i nitrogenfikserende mikroorganismer og består av et jernprotein og et større molybden-jernprotein. ATP kravet er stort, vanligvis fra 20-30 mol ATP per mol fiksert nitrogen. Protonering av molybden er første trinnet i nitrogenfikseringen og da kan hydrogen frigis. Opptil 25 % av elektronstrømmen gennom nitrogenase kan brukes til å lage hydrogen som blir et biprodukt.
Effektiviteten til nitrogenfikseringen avhenger av i hvilken grad bakteriene klarer å ta hånd om utskilt hydrogen og refiksere det med en hydrogenopptaks hydrogenase.
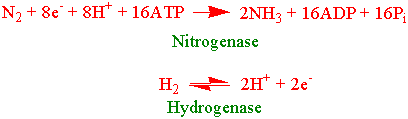
Nitrogenase kan også bruke acetylen som substrat og produktet blir etylen. Bruk av acetylen som kunstig substrat er en viktig metode for å måle nitrogenaseaktivitet. Elektronene til nitrogenfikseringen kommer fra redusert ferredoksin. Nitrogenase er ekstremt følsom for oksygen, og nitrogenfikserende organismer har valgt forskjellige strategier for å kontrollere og holde lav oksygenkonsentrasjon i nærheten av nitrogenase. Bakterier som lever under anaerobe forhold f.eks. Clostridium har løst problemet ved at de vokser på steder uten oksygen.
Bakterier kan også lage mikroaerofile omgivelser ved å bruke opp oksygen til respirasjon, altså respiratorisk beskyttelse av nitrogenase f.eks. Azotobacter. Gloeotheca utfører fotosyntese om dagen og nitrogenfiksering om natten. Nitrogenfikserende bakterier kan også leve i kolonier med en slimskjede som begrenser diffusjonen av oksygen. Nitrogenase kan beskyttes av mekaniske barrierer. Blågrønnbakterier som vokser i miljø med liten tilgang på ammonium f.eks. Anabaena har fysisk atskilt nitrogenase i heterocyster med hard vegg. Heterocystene inneholder bare fotosystem I slik at fotosyntesen utvikler ikke oksygen i heterocystene. Blågrønnbakterier som mangler heterocyster kan bare fiksere nitrogen under anaerobe betingelser. Nitrogenfikserende bakterier i symbiose med erteplanter kontrollerer oksygentilgangen via proteinet leghemoglobin i rotknollene. Leghemoglobin ligner på hemoglobin vi har i blodet og har fått navnet leg fra Leguminosae, erteblomstfamilien. Det har en rustbrun farge som kan observeres hvis man knuser de nitrogenfikserende rotknollene mellom hendene. Bakteriene lager hem og plantene proteinet.
Gras kan dannes assosiasjoner (pseudosymbiotisk) med frittlevende nitrogenfikserende bakterier i slekten Azospirillum, men disse danner ikke nitrogenfikserende knoller. Actinomyceter fra slekten Frankia danner nitrogenfikserende symbiose med røttene på or og pors.
Nitrogenfiksering hos planter og rhizobia
Bakterier fra slektene Rhizobium leguminosarum med kultivarene phaseoli, trifolii og viciae inngår i nitrogenfikserende assosiasjon med henholdsvis bønner, kløver og erter. Bradyrhizobium kan inngå i assosiasjon med Parasponia som ikke er en erteplante og Bradyrhizobium japonicum og Sinorhizobium fredii fikserer nitrogen hos sojabønne. Azorhizobium danner symbiose med vannplanten Sesbania og Photorhizobium danner symbiose med vannplanten Aeschenomene. Alle bakteriene nevnt ovenfor samles under fellesnavnet rhizobia.
Første trinn i infeksjonen er at mikrosymbionten kjenner igjen planten. Flavonoider f.eks lutein utskilles fra røttene hos erteplanter og virker i lave konsentrasjoner som et signal for kjemotaksis for frittlevende Rhizobium. Flavonoidene stimulerer uttrykk av noduleringsgener (nod) i Rhizobium ved høyere konsentrasjoner av flavonoider nær jordoverflaten. nod-genene lager et kjemisk motsignal kalt nod-faktorer. Nodfaktorene er signalstoffet lipokitinoligosakkarider som er oligomere av b-1,4- bundet N-acetylgukosamin bundet til en fettsyre. Disse gir krølling av rothår og deling av celler i rotbarken (cortex). Genene i planten som deltar i dannelsen av rotnoduler kalles nodulingener (Nod). Induksjonen av noduleringsgener trengs for produksjon av sukkerbindende proteiner (lektiner) i rothårene og festing av bakteriene til rothårene. Nod-faktorene er morfogener som får planten til å starte et program for dannelse av rotnoduler. Tidlig fase av assosiasjonen mellom bakterie og plante starter med krølling av rothårene. Planten lager en infeksjonstråd som bakteriene kan følge inn i planten. Infeksjonstråden er en utvidelse av plasmamembranen i planten laget av vesikler fra Golgiapparatet. Forskjellige signalstoffer deltar når det dannes et nodulprimordium i rotbarken (cortex). I spesielle nodulceller frigis bakteriene fra infeksjonstråden i plantenes cytoplasma og omgis av en planteavledet membran peribakteroidmembran. I løpet av denne prosessen blir bakteriene bakteroider som er mange ganger større enn bakteriene og mangler cellevegg. En rotcelle kan inneholde opptil 20000 bakterioder. Overgangen fra bakterier til bakterioder er koblet sammen med syntese av leghemoglobin, nirogenase og andre nitrogenfikserende enzymer. Nitrogenfikseringen starter ca. 10-20 dager etter infeksjonensprosessen ble påbegynt. Det blir lav oksygenkonsentrasjon i nodulene på grunn av leghemoglobin, gassdiffusjonsbarriere i cortex til nodulene og høy respirasjonsrate til bakteriodene. Dette gir energi, men elektronoverføringene gir også mulighet for dannelse av skadelige oksygenradikaler. Nodulene inneholder enzymene i askorbat-glutathionsyklus som beskyttelse mot skadelige radikaler. Ammonium dannet under nitrogenfikseringen assimileres via enzymene GS/GOGAT. Assimilert ammonium fraktes opp i planten i form av aminosyrene glutamin og asparagin. Planter med mer tropisk utbredelse frakter også nitrogen i form av ureider. Ureiedene kan være allantoinsyre, allantoin og citrullin.
{kind=link}
Høyt krav til fosfor gjør at mange erteplanter også har vesikulær-arbuskulær mykorrhiza. Kobolt trengs av bakteriene for å lage leghomoglobin. Nikkel trengs i hydrogenopptaks hydrogenase. Større mengder nitrat hemmer aktiviteten til nitrogenase. Glutamin har 2N/5C, asparagin 2N/4C, arginin 4N/6C. , allantoin og allantoinsyre har 4N/4C.
pH-regulering og opptak av nitrat og ammonium
Nitratopptak gir mer basisk rotmiljø (alkalisering) enten ved en H+/NO3- symport eller en OH-/NO3- antiport. Opptak av ammonium gir en forsuring av rotsonen ved utskillelse av protoner i bytte mot ammonium. Calcifuge planter, - planter som er tilpasset sur jord og lavt redokspotensial foretrekker ammonium som N-kilde, selv om de fleste planter vokser best hvis de får både ammonium og nitrat. Calcicole, planter som foretrekker kalk og høy pH i jorda foretrekker nitrat. Ammonium og nitrat står for 80 % av ioneopptaket av mineralnæringsstoffer.