Enzymet er lite effektivt og har liten affinitet for substratet CO2 (Km-verdi 10-20 mM), og har heller ikke maksimal reaksjonshastighet ved dagens atmosfærekonsentrasjon av CO2 (ca. 340 µmol mol-1), noe som gir en konsentrasjon av karbondioksid i cellen på ca. 10 mM. For å kompensere for dette finnes store mengder rubisko i kloroplastene, og kan utgjøre opptil 50% av alt proteinet i et blad. Enzymet kan også bruke oksygen som substrat. I dette tilfellet blir produktene 3-fosfolycersyre og fosfoglykolat. Fosfoglykolat fungerer som substrat i fotorespirasjon. Ineffektiviteten til rubisko skyldes også fotorespirasjon. Aktiviteten til rubisko reguleres indirekte av lys via aktivering med Mg2+ og CO2. CO2 fungerer altså både som substrat og aktivator. Aktiveringen av rubisko skjer via proteinet rubisko aktivase.
Rubisko danner bro mellom uorganisk og organisk karbon
Rubisko (ribulose-1,5-bisfosfat karboksylase-oksygenase) er et enzym i fotosyntesen som katalyserer det første CO2-fikserende trinnet i Calvin-Bensonsyklus. Alt liv er avhengig av karbon og molekylene er laget av et organisk karbonskjelett. Rubisko sørger for at CO2 kommer inn i organiske forbindelser, og fikserer ca. 1011 tonn CO2 hvert år. Enzymet ble oppdaget av Melvin Calvin og medarbeidere på 1940-tallet via bruk av den radioaktive karbonisotopen 14C.
En fem-karbonforbindelse ribulose-1,5-bisfosfat (RuBP) reagerer med CO2. Først fjernes et proton (H+) ved karbonatom tre. CO2 bindes til enediol-intermediatet og via et C6-intermediat (2-karboksy-3-keto-arabinitol,1-5-bisfosfat) som blir hydrolysert og klippet i to like deler dannes to molekyler 3-fosfoglycerssyre (3-PGA). Planter som bruker rubisko som det primære enzymet for fiksering av CO2 kalles C3-planter. C4-planter og CAM-planter bruker i stedet fosfoenolpyruvat karboksylase til primær CO2-fiksering, og deretter brukes rubisko, atskilt i rom og tid for disse to plantetypene.

Calvin-Benson syklus
Calvin-Benson-syklus består av tre faser: 1) Karboksylering og fiksering av CO2 via rubisko. 2) Reduksjon av 3-fosfoglycerat i to enzymatiske trinn til 3-fosfoglyceraldehyd, ved hjelp av ATP og NADPH laget i lysreaksjonen.. 3) Regenerering av CO2-akseptoren RuBP. 5/6 av 3-fosfoglyceraldehyd som blir laget i Calvin-Benson-syklus brukt til å regenere CO2-akseptoren RuBP, 1/6 blir brukt til å lage andre organiske forbindelser.
Oksygenase med O2 som substrat
Rubisko katalyserer også en oksygenasereaksjon mellom oksygen (O2) og RuBP hvor produktet er ett molekyl 3-fosfoglycersyre og ett molekyl med to-karbonforbindelsen 2-fosfoglykolat, hvor sistnevnte danner utgangspunkt for fotorespirasjon hos C3-planter. Oksygen og karbondioksid konkurrerer om samme bindingssete til enzymet. Hos plantene er rubisko lokalisert til stroma i kloroplastene, mens hos bakterier finnes det i små vesikler kalt karboksysomer.
Små og store subenheter
Rubisko type I finnes hos alle eukaryote fotoautotrofe organismer, og er satt sammen av store (L- ”large”) og små (S- ”small”) subenheter. Rubisko type II finnes i bakterier og inneholder bare de store subenhetene som en dimer. Både katalytisk effektivitet og strukturen til rubisko varierer mellom de forskjellige gruppene fotosyntetiserende organismer. Den store subenheten (55 kDa) er kodet av genet rbcL i kloroplasten og inneholder katalyttisk sete og bindingsseter for reguleringsmetabolitter. Den lille subenheten er kodet av genfamilien rbcS i cellekjernen. Den lille subenheten blir laget på ribosomene i cytoplasma, og inkludert signalsekvensen som sørger for at proteinet blir tatt opp av kloroplastene er størrelsen 20 kDa. For å komme inn i kloroplastene må den lille subenheten passere to membraner. Størrelsen blir for den lille subenheten14 kDa når den er prosessert i kloroplasten. De fleste proteiner har hydrofile overflater og må passere en pore eller proteinkanal for å komme gjennom den hydrofobe indre delen av membranene. Import inn i kloroplasten, og deretter pakking og tilpasning av de forskjellige subenhetene krever molekylære chaperoner. Det ferdige rubisko inneholder strukturen (L2)4(S4)2, fire dimere med store subenheter og to dimere med en tetramer av de små subenhetene, altså L8S8 i planter.
Rubisko er det hastighetsbegrensene trinnet i CO2-fikseringen. Enzymet er lite effektivt og katalysere fra ca. 3 molekyler per sekund, mens andre enzymer kan være fra hundre til tusenvis ganger mer effektive. Den manglende effektivitet blir kompensert med tilsvarende økt mengde enzym, og rubisko utgjør opptil halvparten av proteinene man finner i grønne blader.
Aktivering av rubisko
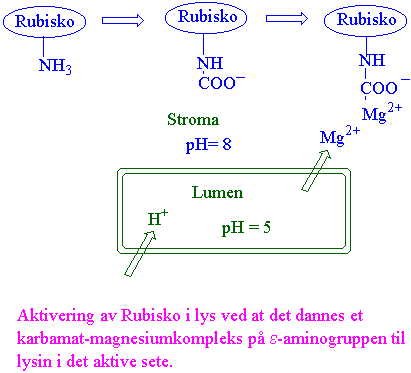
Rubisko er aktivt om dagen og inaktivt i mørke. Aktivering av rubisko skjer ved at et molekyl med aminosyren 201 lysin blir positivt ladet og kan binde CO2 kovalent og denne et karbamylat. CO2 som blir brukt til aktivering er forskjellig fra CO2 som blir fiksert. Det karbamylerte enzymet danner et ternært kompleks med magnesium.
(Enzym-lysin-NH2)inaktiv + CO2 + Mg2+ → (Enzym-lysin-NHCOO-Mg2+)aktiv + H+
Rubisko blir regulert av fosforylerte metabolitter og det hemmende stoffet KA1P (2-karboksy-D-arabinitol-1-fosfat). KA1P blir laget fra 2-karboksy-D-arabinitol som kommer fra fruktose-1,6-bisfosfat, et intermediat i Calvin-Benson-syklus.
KA1P blir bundet til karbamylert rubisko-Mg2+ -kompleks via enzymet rubisko aktivase som reagerer på redoksnivået i kloroplastene. Rubisko aktivase finnes i to former med størrelse 42 kDa og 46 kDa. KA1P blir fjernet via en fosfatase.
{kind=link}