
TEM-bilde 1 av blad fra ung frøplante ert (Pisum sativum L.) tatt ved Elektronmikroskopisk laboratorium, UiO. Bildet viser overgangen mellom to celler atskilt av cellevegg og midtlamell med kloroplaster på hver side av celleveggen. Kloroplaster (K) er omgitt av dobbel membran. Inne i kloroplastene er det tylakoidmembraner hvor lysreaksjonen foregår (grana og intergranalameller), og som virker som ankerfeste for lyshøstende protein-pigmentkomplekser (klorofyll, karotenoider), elektrontransportenzymer og ATPsyntase) . Mellom og rundt tylakoidmembranene er stroma med assimilasjon av karbondioksid i Calvin-syklus . Bildet viser mitokondrier (M), plasmamembranen inn mot celleveggen og tonoplast (vakuolemembranen) mot vakuolen som sees lengst til venstre i bildet. Cytosol inneholder vesikler fra Golgi-apparatet og endoplasmatisk retikulum. Vi kan også se to stivelseskorn inne i den nederste kloroplasten. Stivelse dannes bare i plastider, det vil si her inne i kloroplasten. Kloroplastene er fra 5-10 µm lange. Målestokk 1 µm.
Kloroplasten kan også inneholde stivelseskorn og fettdråper (plastoglobuli). Plastoglobuli innholder lipoprotein, er festet til tylakoidmembranene, og produserer tokoferoler (vitamin E) som deltar i antioksidantforsvaret og fjerner oksygenradikaler. Plastoglobuli deltar også i produksjonen av fyllokinon (vitamin K1).
Det er ikke bare enzymsystemene for fotosyntese som befinner seg i kloroplasten. Enzymer for assimilasjon av ammonium og biosyntese av aminosyrer, samt biosyntese av fettsyrer er lokalisert til kloroplasten. Kloroplastene inneholder mye lipider av typen monoglaktosyldiacylglycerol (MDDG) og digalaktosyldiacylglycerol, og mindre mengder fosfatidylglycerol (PGH) og sulfoquinovosyldiacylglycerol (SQDG)- Kloroplasten inneholder et sirkulært DNA og 70 S ribosomer. Kloroplaster lages fra proplastider. Algekloroplaster har ikke grana.

TEM-bilde 2 av blad fra ung frøplante ert (Pisum sativum L.) tatt ved Elektronmikroskopisk laboratorium, UiO. Bildet viser kloroplaster, mitokondrier og en cellekjerne som ligger i cytplasma, avgrenset av plasmamembranen mot celleveggen og vakuolemembranen (tonoplasten) mot vakuolen til høyre i bildet. I cellekjernen kan man observere heterokromatin, eukromatin og nukleolus. Målestokkmarkør 2 µm.

TEM-bilde 3 av blad frøplante ert (Pisum sativum L.) tatt ved Elektronmikroskopisk laboratorium, UiO. Bildet viser to celler som ligger inntil hverandre atskilt av cellevegger. Lengst til høyre og venstre i bildet er vakuolene fra de to cellene. Bildet viser kloroplaster med tylekoidmembraner, mitokondrier med mitokondriemembraner (cristae) og en cellekjerne med eukromatin og heterokromatin. Målestokkmarkør 2 µm

TEM-bilde 4 av blad frøplante ert (Pisum sativum L.) tatt ved Elektronmikroskopisk laboratorium, UiO. Bildet viser planteceller med kloroplaster, mitokondrier og cellekjerner. Legg merke til at kloroplastene ligger ut mot celleveggen. Diffusjon av gasser i bladet skjer i intercellularrommene, og det blir forholdsvis kort vei for CO2 å diffundere gjennom celleveggen, gjennom plasmamembranen via akvaporiner, og videre gjennom cytoplasma før CO2 kommer fra til kloroplastene og gjennom kloroplastmembranene via kloroplastakvaporiner. Gassdiffusjon skjer ca. 10000 ganger raskere i gassfase enn i væskefase, og ifølge Ficks andre diffusjonslov skjer diffusjonen raskt over korte avstander, som her. Imidlertid tar diffusjon over store avstander ifølge Ficks andre lov svært lang tid. Målestokkmarkør 10 µm

TEM-bilde 5 av blad frøplante ert (Pisum sativum L.) tatt ved Elektronmikroskopisk laboratorium, UiO. Bildet viser flere fotosyntetiserende celler i bladet. Uenfor celleveggene er det intercellularrom med gassdiffusjon. Evolusjon av store vakuoler i planter gjorde det mulig for plantene å øke i størrelse uten at det medførte for store kostnader i investering av ressurser. Ficks diffusjonslover er viktig for å kunne forklare hvorfor et blad er konstruert slik det er. Målestokkmarkør 10 µm
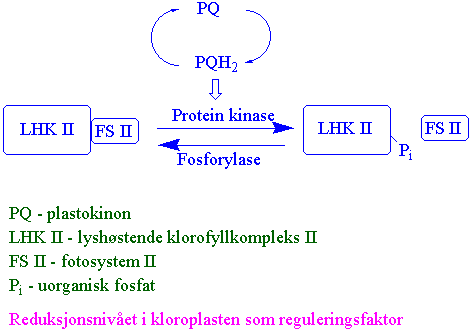
Reduksjonsnivået i kloroplasten som reguleringsfaktor
{kind=link}
Encellete alger inneholder vanligvis en kloroplast per celle. Formen på den plateformete kloroplasten hos Mougeotia påvirkes av rødt og mørkerødt lys via fytokromsystemet. Kloroplastene festet til cytoskjelettet og motorproteiner kan forflytte seg i plantecellen avhengig av lysfoholdene i bladet. Blått lys absorbert av kryptokrom deltar i denne forflytningen.
Kloroplaster utviklet hos planter som ikke kan lage klorofyll i mørke kalles etioplaster, med et semikrystallinske membransystem (prolammelarlegemer) og protoklorofyll.. Andre typer plastider er kromoplaster som inneholder karotenoider, og amyloplaster som inneholder stivelse. Disse kan omdannes til hverandre. Kloroplaster omdannes til kromoplaster i frukt og høstblad. I poteter som ligger i lys omdannes hvite amyloplaster til grønne kloroplaster. Når tomater modner omdannes kloroplastene til kromoplaster med stort innhold av karotenoidet lykopen. Grønne kloroplaster i bananer omdannes til gule kromoplaster når bananen modner. De fleste fotosynteseenzymer er kodet av kjernen og importert.
Kloroplaster er semiautonome organeller som inneholder kloroplast-DNA (kp-DNA) ca. 145 kbp (kilobasepar), og det er polyploidi for kp-DNA i en kloroplast. Mitokondrie-DNA (mt-DNA) og kp-DNA utgjør ca. 1/3 av kjernegenomet. Fra kp-DNA lages ribosomalt RNA(rRNA), transport RNA (tRNA), og bla. annet den store subenheten til rubisko.De fleste fotosynteseenzymer er imidlertid kodet av cellekjernen og importert merket med signalpeptider. Inne i kloroplasten pakkes ved hjelp av chaperoner flere enheter med den lille subenheten til rubisko kodet fra cellekjernen sammen med store subenheter laget i kloroplasten. Sirkulært DNA er lokalisert i deler av stroma i et matriks kalt nukleoider. Replikasjon i kloroplaster skjer uavhengig av cellekjernen. Plastider forbundet med stromafylte rør danner et nettverk av plastider kalt stromuler.
Begge de ytre to kloroplastmembraner inneholder mye glykosylglycerider (galaktoglycerolipider) med prokaryot opprinnelse, og den indre av de ytre membranene har samme innhold som og de indre tylakoidmembraner .
Isolering av kloroplaster
Spinat (Spinacea oleracea) er den planten hvor det er lettest å isolere intakte kloroplaster. Andre planter som er mulig å prøve er ert (Pisum sativum), salat og meldestokk (Chenopodium album).
Blader avkjøles, klippes og knuses i et isotont medium i en blender i ca. 20 sekunder. Ekstraktet filtreres gjennom gaz og nettingduk. Etter sentrifugering f.eks. 1.000 xg i 10 minutter ristes pelleten forsiktig opp i det isotone mediet. Disse kloroplastene kan brukes til å studere Hill-reaksjonen i en fargereaksjon med 2,6-diklorfenol-indofenol. Kloroplaster kan renses ytterligere via gradientsentrifugering