Ryggstrengdyrene (chordatene) har en langsgående dorsal brusklignende ryggstreng (notochord, notokord; gr. noton - rygg; chorde - streng; chorda dorsalis) som skjelettstreng/støttestruktur/skjelettstav, ihvertfall i embryo/larvestadiet, og som ligger dorsalt for urtarmen, det vil si mellom fordøyelseskanal og nerverøret.
Ryggstrengen, som ikke må forveksles med nervestrengen, er en relativt stiv fleksibel stavformet struktur med store væskefylte celler som støtter kroppen, som går fra spiss til hale, og er omgitt av en fiberskjede og ytterst en elastisk skjede. Lateralt er ryggstrengen fleksibel og gir mer effektiv svømming. Ryggstrengen gir feste for muskler. Kroppen er segmentert, selv om det ikke alltid er like lett synlig. Hos Protochordater og kjeveløse fisk (niøyer og slimål) er ryggstrengen varig, men hos de fleste vertebratene blir den embryonale ryggstrengen erstattet med en mer kompleks virvelsøyle (vertebra; l. vertebra - ryggvirvel). Rester av ryggstrengen finnes som gelatinformede skiver mellom eller i ryggvirvlene. Ryggvirvlene hos vertebratene dannes fra mesenkymceller avledet fra grupper av mesodermale celler på siden av ryggstrengen.
Ryggstrengdyrene har endoskjelett, og lukket sirkulasjonssystem med et ventralt plassert hjerte. Kroppen er utformet som et rør i et rør. Dorsalt for fordøyelseskanalen og ryggstrengen ligger hos chordadyrene en hul rørformet nervestreng (nerverør) dannet i embryo fra en innfolding og fortykning i ektodermen. Nervestrengen danner hjerne og sentralnervesystem. Hos leddormer (annelider) og leddyr (artropoder) er nervestrengen ikke hul og den ligger ventralt plassert i forhold til fordøyelseskanalen. I framenden er nervestrengen utvidet og danner en hjerne beskyttet av et kranium. Hos vertebratene ligger nervestrengen beskyttet i nervebuer i ryggsøylen. Chordatene har samme fellestrekk som deuterostome dyr (Deuterostomia): radial kløyving og første embryonalåpning/urmunnen i blastoforen blir til analåpning. Coelomet er enterocoel, har tre velutviklede kimlag, og den sekundære kroppshulen utvikles fra mesodermen. Ryggstrengdyrene har kjeveposer i minst ett av stadiene i livssyklus, og utvider seg fra fremre del av fordøyelsestrakten mot gjellespaltene.
For arter som lever i vann er veggen i svelget gjennombrutt av gjellespalter som benyttes til respirasjon og filtrering av næring. Vann som kommer inn i munnen kan gå ut gjennom gjellespaltene. Opprinnelig fungerte gjellespaltene til filterspising, men fikk under evolusjonen betydning i gassutvekslingen. Gjellestavene gir støtte for kjever og deltar i utvikling av hørselorganer. Vann og næringspartikler kommer inn gjennom munnen og gjellefolder med slim fanger fødepartikler. I stedet for å gå gjennom tarm og anus føres vannet ut gjennom gjellespaltene til utsiden. Det perforerte svelget hos protochordatene virker som et filterapparat, hvor vannet føres inn i munnen med cilibevegelser. Hos vertebratene er ciliene erstattet av muskler som driver vannet gjennom svelget ved å utvide og trekke sammen hulrommet i svelget. I samvirke med utviklingen av aortabuene blir gjennom evolusjonen filtreringsapparatet hos prochordatene til indre gjeller som brukes til respirasjon hos de akvatiske vertebratene. Det er gjellespalter i kroppsveggen i svelget i en del av livssyklus. Landlevende krypdyr og fugler har de alternerende gjellespaltene og gjellebuene bare i fosterstadiet. De første ryggstrengdyrene var suspensjonsspisere, og etter hvert utviklet fremre gjellebue seg til kjever. Hos tetrapoder deltok kjeveposene i utviklingen av hulrommet i mellomøret, øretrompeten (eustachiske rør), mandler, skjoldbruskkjertel. Hjertet er ventralt plassert i forhold til ryggstreng og tarm. Hos chordatene, som har lukket blodkarsystem, ligger hovedblodåren ventralt og blodet beveger seg forover ventralt.
Ryggstrengdyrene har en muskulær postanal hale (l. post - etter; natalis - fødsel) som vanligvis stikker ut bak anus, og har på samme måte som ryggstrengen en viktig funksjon ved bevegelse av kroppen. Hos mennesker er det bare rester av halen som et halebein Ryggstrengdyrene har et fullstendig fordøyelsessystem. Musklene er segmenterte.
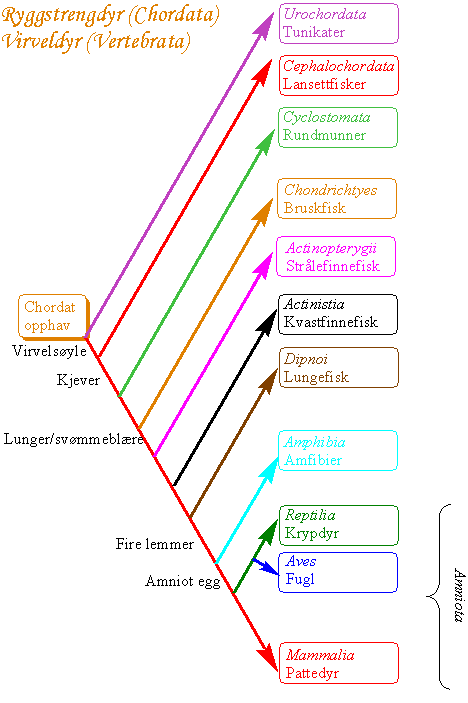
Virveldyrene (vertebratene) har et skjelett bestående av brusk og/eller bein, og har en skalle som omgir hjernen og har blitt plassert i kraniedyrene (Craniata). Kraniedyrene (Craniata) som omfatter vertebratene og slimål (Myxinoidea), består av tetrapoder og superklassen fisk (Pisces) med niøyer (Cephalaspidomorphi) og fire klasser kjevefisk: (actinistier som omfatter rhipidistier og kvastfinnefisk (Actinistia), strålefinnete fisk (Actinopterygii), bruskfisk (Chondrichthyes) og lungefisk (Dipnoi)).
Tetrapodene omfatter amfibier (Amphibia), og de amniote krypdyr (Reptilia), fugl (Aves) og pattedyr (Mammalia). Tetrapodene har fire hox-klynger, hver med 10 gener, men sebrafisk har 7 hox-klynger.
Protochordata mangler skalle og hører med til Acraniata. Grupperingene kjeveløse virveldyr (Agnatha) og krypdyr (Reptilia) er parafyletiske og tilfredsstiller ikke kravet om at taxa skal være monofyletiske. Imidlertid hører både krypdyr, fugl og pattedyr til den monofyletiske Amniota, dvs. de utvikles fra et egg med spesielle fosterhinner bl.a. den innerste fosterhinnen amnion. Vertebratene fisk og amfibier som mangler amniote egg hører med til Anamniota. Virveldyr med kjever ble i det linneiske system plassert i Gnathostomata. Det er evolusjonært slektskap mellom primitive pigghuder og chordatene utviklet fra et felles deuterostomt opphav. Fossile pigghuder i Calcichordata hadde gjellespalter, men i motsetning til chordatene hadde de et skjelett av kalsiumkarbonat. Deuterostome dyr (pigghuder, hemichordater, lophophorater og chordadyr) hadde felles opphav i prekambrium. Primitive chordate fossiler er Pikaia funnet i Burgess Shale i Canada, som ligner lansettfisk .
Det er omdiskutert om fossile Yunnanozoer f.eks. den ca. 3 cm lange Haikouella fra Kina var deuterostome ryggstrengdyr eller ikke. Haikouella hadde hjerne, men manglet kranium. Den hadde også øyne i motsetning til lansettfisker og hadde også tannlignende strukturer i svelget En annen Yunnanzoo, den 2-3 cam lange Haikouichthys hadde kranium og flere utviklingstrekk som vertebratene. Haikouichthys og den nærstående arten Myllokunmingia er blant de første vertebratene. De var sannsynligvis predatorer, det vil si ikke suspensjonsspisere som lansettfiskene. Det betyr at vertebratene har sin opprinnelse fra den kambriske eksplosjon. Tarmsjøpung (Ciona intestinalis) har enkle former av vertebrate genfamilier som viser slektskapet.
Teksten er hentet fra Deuterostomia