Fotosyntese
Assimilasjon av karbondioksid
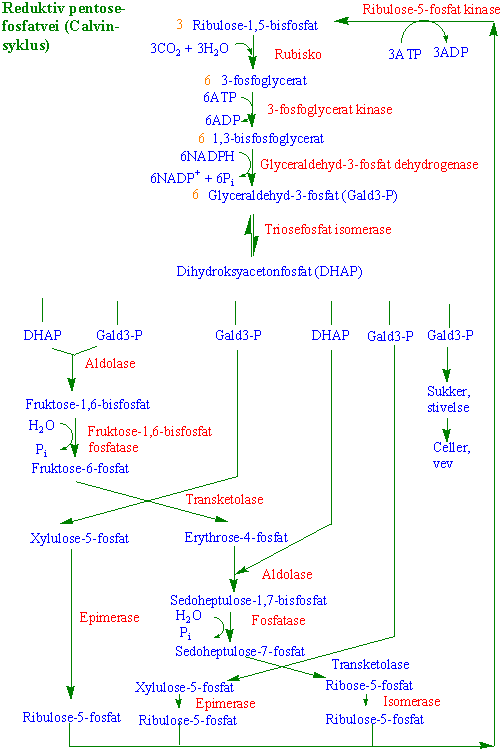
- Reduktiv pentosefosfatvei (Calvin-syklus)
- C4-metabolisme
- CAM-metabolisme
- Fotorespirasjon
- Åpning og lukking av spalteåpninger
- Fotosyntese hos lys- og skyggeplanter
- Oppkonsentrering av karbondioksid
- Fosfattranslokator
For å kunne utnytte energien som er lagret i lysreaksjonen i form av ATPog NADPH, trengs en CO2-akseptor før CO2 kan bli redusert: ribulosebisfosfat - en 5-karbonforbindelse. I to trinn blir 3- fosfoglycersyre redusert ved hjelp av ATP og NADPH til glyceraldehyd-3-fosfat (triosefosfat). CO2-akseptoren ribulosebisfosfat regenereres via Calvin-Benson-syklus. Resten av triosefosfatene brukes til lage stivelse i kloroplasten eller blir transporteret som C3-forbindelser ut av kloroplasten (Figur). Mengden av C3 som fraktes ut av kloroplasten kontrolleres av tilgangen på uorganisk fosfat (Pi). CO2 blir ikke bare assimileret via C3-fotosyntese som beskrevet over. CO2 kan også assimileres via fosfoenolpyruvat-karboksylase oppdaget av Kortschack, Hatch og Slack. Fosfoenolpyruvat fungerer som CO2- akseptor. C4 planter er arter med tropisk opprinnelse. C4-fotosyntesen gjør at plantene kan utnytte CO2 og vann mer effektivt enn C3-plantene. Dette skyldes at fosfoenolpyruvat karboksylase har høy affinitet for CO2 .
{kind=link}
Reduktiv pentosefosfatvei (Calvin syklus)
Alle planter og alger kan redusere CO2 til karbohydrater ved samme mekanisme kalt reduktiv pentosefosfatvei eller Calvinsyklus. Denne består av 13 enzymkatalysert reaksjoner i stroma i kloroplastene. Fotosyntetisk fiksering og assimilering av karbondioksid i kloroplaster ble vist første gang av Arnon i 1954. Melvin Calvin og medarbeiderne Benson og Bassham klarla på 1950-tallet hvordan CO2 ble assimilert i grønnalgen Chlorella. Ved hjelp av 14C, todimensjonal kromatografering og autoradiografi utledet de sekvensen av reaksjoner. Calvin fikk nobelprisen i kjemi i 1961 for dette arbeidet.
{kind=link}
Syklusen består av tre hovedtrinn
{kind=link}
- 1) Karboksylering
- 2) Reduksjon
- 3) Regenerering
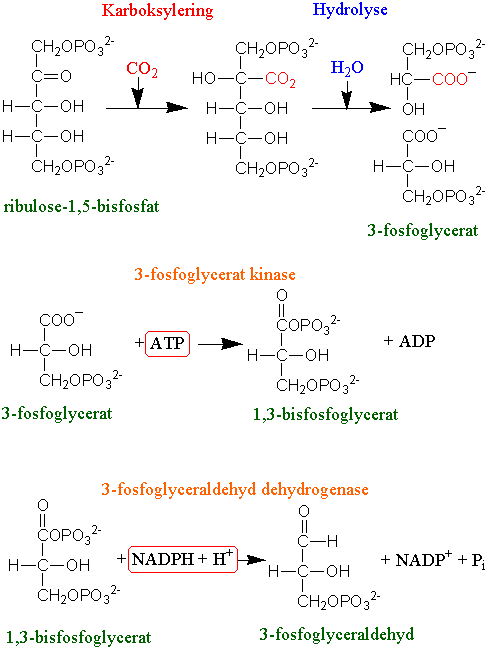
Karboksyleringen skjer ved at fem-karbon akseptoren ribulose 1,5-bisfosfat (RuBP) bindes til CO2 og vann katalysert av enzymet ribulose 1,5-bisfosfatkarboksylase/oksygenase med akronymet rubisko. Oksygenaseaktiviteten til rubisko gir opphav til fotorespirasjon.
{kind=link}

Opptil 40% av det løselige proteinet i blader består av rubisko. Rubisko er satt sammen av 8 store subenheter kodet av kloroplastgenomet og 8 små subenheter kodet fra cellekjernen (L8S8). Total molekylvekt er ca. 560 kDa i en konsetrasjon tilsvarende ca. 4 mM. Den lille subenheten er merket med en transit peptid i N-enden som anviser at den skal til gjennom kloroplastmembranen. Sammenkoblingen av subenhetene i rubisko til riktig konformasjon er katalysert av chaperoner.
Ved hydrolyse dannes det to molekyler 3-fosfoglycersyre som er de første stabile intermediatene i syklusen. Reduksjonen skjer ved at 3-fosfoglycerat fosforyleres til 1,3-bisfosfatglycerat ved hjelp av ATP fra lysreaksjonen katalysert av enzymet 3-fosfoglycerat kinase. Vi bruker endelsen -at når det er snakk om den ioniserte syren og -syre når den er protonisert.

NADPH brukes deretter til å lage 3-fosfoglyceraldehyd katalysert av NADP-glyceraldehyd-3- fosfat dehydrogenase

Det samme enzymet i glykolysen bruker NAD som koenzym. 3-fosfoglyceraldehyd kan omdannes til dihydroksyacetonfosfat i en isomeringsreaksjon katalysert av triosefosfat isomerase.

Glyceraldehyd-3-fosfat og dihydroksyacetonfosfat kalles triosefosfater. Triosefosfater eksporteres ut av kloroplasten i bytte med ortofosfat via en triosefosfat translokator. 5 molekyler triosefosfat ut av 6 nylagete trengs for å regenerere RuBP. For hver av de 6 molekyler triosefosfat trengs 5 for å regenerere tre molekyler ribulose 1,5-bisfosfat. Dihydroksyacetonfosfat og glyceraldehyd-3-fosfat gjennomgår en aldolkondensasjon katalysert av en aldolase og produktet er fruktose-1,6-bisfosfat

Fruktose-1,6-bisfosfat hydrolyseres til fruktose-6-fosfat katalysert av fruktosebisfosfat fosfatase
![]()
En enhet på to karbonatomer overføres fra fruktose-6-fosfat til et tredje molekyl 3- fosfoglyceraldehyd og gir erythrose-4-fosfat og xylulose-5-fosfat katalysert av en transketolase.

Erythrose-4-fosfat kombineres med et fjerde molekyl 3-fosfoglyceraldehyd og gir sjukarbonsukkeret sedoheptulose-7-fosfat katalysert av en aldolase

Sedoheptulose-1,7-bisfosfat hydrolyseres til seduheptolose-7-fosfat katalysert av sedoheptulose-1,7- bisfosfatase
![]()
Sedoheptolose avgir to karbonatomer til et femte og siste molekyl glyceraldehyd-3-fosfat, katalysert av en transketolase og produktene er xylulose-5-fosfat og ribose-5-fosfat.

De to molekylene med xylulose-5-fosfat omdannes til to molekyler ribulose-5-fosfat katalysert av en epimerase
![]()
og ribose-5-fosfat omdannes til ribulose-5-fosfat katalysert av en isomerase.
![]()
Ribulose-5-fosfat kinase fosforylerer de 3 molekylene med ribulose-5-fosfat til 3 molekyler med CO2-akseptoren ribulose-1,5-bisfofat.
![]()
I sammendrag deltar de følgende enzymer i de 13 reaksjonene:
- 1. Ribulose-bisfosfat karboksylase
- 2. Fosfoglycerat kinase
- 3. NADP-glyceraldehydfosfat dehydrogenase
- 4. triosefosfat isomerase
- 5. Aldolase
- 6. Fruktose bisfosfatase
- 7. Transketolase
- 8. Aldolase
- 9. Sedoheptulose-1,7-bisfosfatase
- 10. Transketolase
- 11. Ribose-5-fosfat isomerase
- 12. Ribulose-5-fosfat epimerase
- 13. Fosforibulokinase.
Reduktiv pentosefosfatvei lager sine egne substrater. Hastigheten på syklusen øker til en maksimal fluks. 1/6 av karbonet er tilgjengelig for eksport. 5/6 for regenerering av RuBP. Strukturformler for intermediater i reduktiv pentosefosfatvei.
{kind=link}
For å lage en heksose fra 6 molekyler med CO2 trengs 18 ATP og 12 NADPH, dvs. det forbrukes 3 molekyler ATP og 2 molekyler NADPH for hvert CO2 molekyl som blir fiksert.
{kind=link}
Calvinsyklus er en energieffektiv prosess. Heksose som blir fullstendig oksidert ved respirasjon frigir 2804 kJ mol-1. Hydrolyse av ATP frigir 29 kJ mol-1 og NADPH som blir oksidert frigir 217 kJ mol-1. Totalt gir dette en effektivitet på ca. 90% (12×217 + 18×29 = 3126 kJ mol-1).
Anaerobe fotosyntetiske bakterier bruker ikke Calvin-syklus. Grønne svovelbakterier kan bruke reduktiv sitronsyresyklus (reduktiv Krebssyklus eller trikarboksylsyresyklus) til å lage organiske syrer som oksalacetat hvor acetyl-CoA brukes som CO2 akseptor. Redusert ferredoksin brukes som elektronkilde og produktet er pyruvat. CO2 kan deretter bindes i overgangen fosfoenolpyruvat til oksalacetat, mellom succinyl-CoA og 2-oksoglutarat og mellom 2-oksoglutarat og oksalosuccinat.
Metanogene og acetogene bakterier kan bruke acetyl-CoA biosynteseveien.
Grønne ikke-svovelbakterier kan bruke hydroksypropionsyre biosynteseveien.
I fullt sollys kan reduktiv pentosefosfatvei begrense fotosyntesen. Feedbackregulering av stroma og thylakoidreaksjoner via tilgjengeligheten av NADPH. Mengden NADP/NADPH i kloroplasten er ca. 20 nmol mg klorofyll-1, hvorav ca. 50 % er i redusert form i lys. NADP+ kan lages fra NAD+ i stroma. Mengden adenylater (ATP, ADP, AMP) er ca. 60 nmol per mg klorofyll
Proteinkonsentrasjonen av rubisko 300 mg ml-1 CO2 konsentrasjonen i kloroplasten ved 25 oC er ca. 10 M ved steady state fotosyntese mens Km for CO2 i luft er ca. 20 mmM. Maksimal karboksylering blir da 1 mg CO2 /h mg karboksylase. Netto fotosyntese for C3 plante er ca. 25 mg CO2 dm-2 h, dvs. det trengs 25 mg RuBP på dette arealet. Den høye Km-verdien for rubisko vil si at med dagens CO2- konsentrasjon går rubisko med halv hastighet. Tilførsel av karbondioksid til C3-planter vil derved gi midlertidig økt vekst hvis næringstilgangen er god. Dette er et fenomen som kalles CO2-gjødsling og er benyttet i veksthusnæringen. Den spesifikke aktiviteten er også lav 1-4 mmol min-1 mg protein-1. Opptil 50 % av bladproteinet kan være rubisko.
Skyggeblad har lavere lysmettet fotosyntese og mindre mengde rubisko enn solblad. Rubisko er fritt løselig i stroma men er bundet med elektrostatiske krefter til thylakoid og kloroplastmembranen. I blågrønnbakterier mangler rubisko i de nitrogenfikserende heterocystene. Dette fordi nitrogenase ikke tåler oksygen. I fotosyntetiserende vegetative celler er rubisko fordelt mellom løselig i cytoplasma og partikulært i karboksysomer.
Regulering av Calvin-syklus
Reduktiv pentosefosfatvei reguleres av lys. Det skjer ingen CO2 assimilasjon i mørke i reduktiv pentosefosfatvei og følgende 5 enzymer i Calvinsyklus reguleres av lys: rubisko, NADPH- glyceraldehyd-3-fosfat dehydrogenase, fruktose-1,6-bisfosfat fosfatase, seduheptolose-1,7-bisfosfat fosfatase og ribulose-5-fosfat kinase. Lyskontrollen skjer bla.a. ved ferredoksin-thioredoksin.
Aktiveringen skjer ved at elektroner fraktes fra redusert ferredoksin til svovelproteinet thioredoksin katalysert av ferredoksin-thioredoksin reduktase. Ferredoksin inneholder to jernatomer og to svovelatomer per molekyl. Ferredoksin kan overføre elektroner til NADP+, glutamat syntetase, nitritt reduktase, thioredoksin og sulfitt reduktase.
Thioredoksin er proteiner med aktive disulfidgrupper og ferredoksin eller NADPH brukes til å lage thioredoksin i redusert form som igjen kan redusere disulfidbindinger i andre proteiner. Hos C4- planter aktiveres NADP-malat dehydrogenase av thioredoksin.
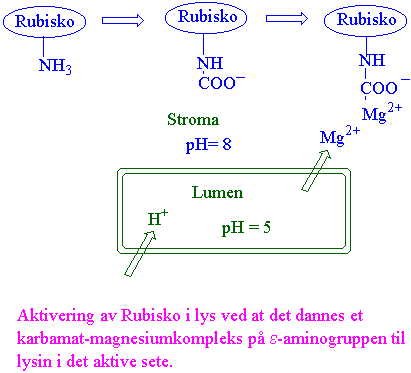
Rubisko aktiveres også av Mg2+ og CO2 hvor CO2 reagerer med e-NH2 på et lysin i det aktive sete i en karbamyleringsreaksjon. Deretter bindes Mg2+. Rubisko aktivase er et enzym som fjerner ribulose-1,5-bisfosfat fra en dekarbamylert inaktiv form av rubisko ved hjelp av ATP, og gir konformasjonsendring til aktiv form av rubisko. Karboksyarabinitol hemmer rubisko og det er mest karboksyarabinitol om natten.
{kind=link}
Reduksjonsnivået i kloroplasten er en viktig reguleringsfaktor i fotosyntesen.
Styres bl.a. i hvilken grad poolen av plastokinon er redusert.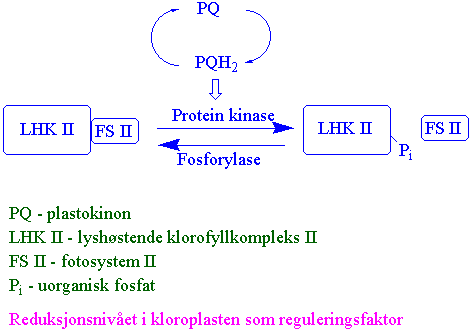{kind=link}
Fotosyntese hos lys- og skyggeplanter
En tett bestand av planter gir en betydelig skyggeeffekt. Ved skrå belysning og med lite optimal plassering av bladene skal det mye til at fotosyntesen er mettet med lys for alle bladene på en plante. Fotosyntesehastigheten er avhengig av lysintensiteten, det vil si hvor mange lyspartikler klorofyllmolekylene kan ta opp. I mørke og svakt lys dominerer respirasjonen (celleåndingen). Etterhvert som lysintensiteten øker stiger fotosyntesen inntil maksimum. En skyggeplante når metning av fotosyntesen ved en lavere lysfluks enn en solplante. Lysintensiteten hvor åndingen og fotosyntesen oppveier hverandre, kalt lyskompensasjonspunktet, varierer fra skyggeplanter til solplanter og er ved henholdsvis ca. 0.5 % og 5 % av fullt sollys. Grunnen til forskjellen er at skyggeplantene har lavere respirasjon enn solplantene. Fotosyntesen hos skyggeplantene er mettet ved ca. 5% av fullt sollys og for solplantene ved ca. 60 %.
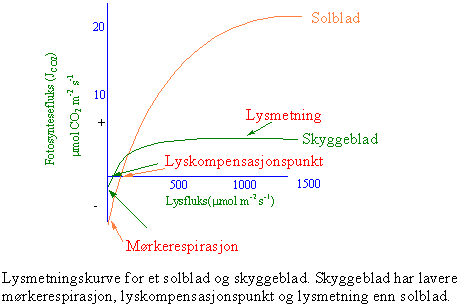
I en tett skog har solglimt gjennom løvverket ned til skogbunnen har stor betydning for skyggeplantene som vokser der, spesielt i tropisk regnskog. Opptil 30-60 % av fotosyntesen skjer i lysglimtene.
Fotosyntesen påvirkes også av konsentrasjonen av karbondioksid. Ved dagens konsentrasjon er ikke fotosyntesen hos C3-planter ikke mettet med karbondioksid. Det er bare ved lite lys at det er nok karbondioksid til at fotosyntesen er mettet

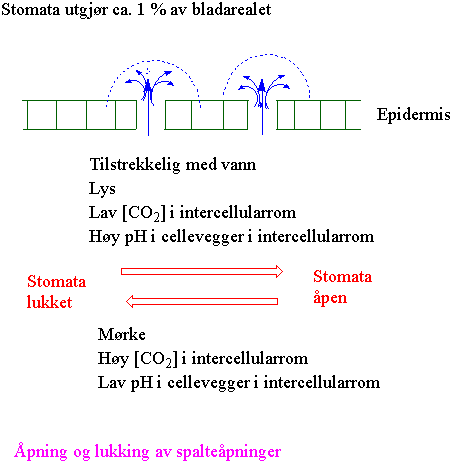
Åpning og lukking av spalteåpninger
På blader og stengler finnes spalteåpninger som virker som regulerbare ventiler. De ligner på små munnåpninger og akkurat som en munn, kan de åpne og lukke seg. Lukkecellene i spalteåpningen er grønne i motsetning til andre celler i epidermis.
Lukkecellene er konstruert slik at spalten lukker seg når lukkecellene mister vann, og åpner seg når de tar opp vann. Grunnstoffet kalium deltar i denne vanntransporten. Går kalium ut av lukkecellene og inn i nabocellene følger vann etter og veggenes form gjør at spalten lukker seg. Går kalium fra nabocellen inn i lukkecellene kommer vann inn og spalten åpner seg.
{kind=link}

Spalteåpningenes areal er bare 1-2 % av det totale bladarealet, men det er allikevel store mengder vann som fordamper ut av disse. Omtrent 90 % av plantenes vanntap skjer via spalteåpningene. De resterende 10 % går igjennom kutikula. Karbondioksid som plantene trenger i fotosyntesen for å kunne vokse kommer gjennom spalteåpningene. Får planten for lite vann fra jorda lages plantehormonet abscisinsyre og spalteåpningene lukkes. Hvis fotosyntesen stopper opp for en tid gjør det ingen skade på kort sikt. Lukkingen av spalteåpningene gir større reduksjon i vanntapet enn den påvirker transporten av karbondioksid inn i bladet. Nok oksygen får plantene selv om spalten er lukket siden det er nesten 600 ganger så mye oksygen som karbondioksid i lufta.
Ved god vanntilførsel er konsentrasjonen av karbondioksid inne i bladet viktig for åpning og lukking av spalteåpningen. Ved høy konsentrasjon, som betyr at fotosyntesen har rikelig tilgang på karbondioksid, lukkes spalten. Ved lav konsentrasjon åpnes spalten for å få mer karbondioksid til plantens fotosyntese. Dette betyr i praksis at spalten er åpen i lys, hvor blått lys er mest effektivt til å lage åpning, og lukket i mørke. Både karbondioksid og vann beveger seg alltid fra det stedet hvor det er høyest konsentrasjon til dit hvor det er lavest. Under aktiv fotosyntese betyr dette at vann går ut av bladet til luften omkring og karbondioksid går inn i bladet. I vindstille vær kan det dannes et lag med stabil luft rundt spalteåpningene slik at luftfuktigheten her øker. Dette gir mindre vanntap. Planten kan redusere vanntapet ved å plassere lukkecellene nedsenket i forhold til epidermis eller ved å skape luftstille rom rundt spalteåpningene ved hjelp av hår. Gras minsker vanntapet ved å rulle sammen bladet.
Sukkulenter som vokser i tørre, varme strøk åpner spalteåpningene om natten og binder karbondioksid fra lufta inn i organiske syrer som lagres i vakuoen. Om dagen når den drepende varmen kommer lukkes spalteåpningene og planten har et lager av karbondioksid inne i planten som den kan benytte på dagtid.
Ved å studere herbariemateriale fra de to siste århundre har det vist seg at antall spalteåpninger blir færre ettersom konsentrasjonen av karbondioksid i lufta øker.
Vanligvis 100-300 stomata per mm2. Hos mjødurt (Filipendula ulmaria) har man funnet 1300 stomata per mm2. Oliventreets blad har ca 600 spalteåpninger per mm2 på undersiden. Et eikeblad har et par millioner spalteåpninger, et middels stort kålblad har 11 millioner (oversiden 400, oversiden 700). Hos gras og bartrær sitter spalteåpningene på lengderekker.
1 g spinatblad inneholder ca 400.000.000 kloroplaster, 1 mg klorofyll, 18 mg kloroplastprotein og har et totalt overflateareal av thylakoider 60 m2
Det finnes ofte stivelseskorn og rundt osmiofile (binder osmium) legemer rike på fett kalt plastoglobuli, inneholder lipider og plastokinin og tokoferylkinon. Kloroplasten inneholder flere kopier av sirkulært DNA.
Fotosyntesen angis som en fluks eller som et kvanteutbytte. Oksygenutvikling kan brukes til å måle ikke-syklisk elektrontransport. Kvanteutbytte minsker når raten for CO2-fiksering øker. Maksimalt utbytte 0.125.
Fosfattranslokator
Fosfat fraktes inn i stroma fra cytosol i bytte med dihydroksyacetonfosfat (triosefosfat). Dihydroksyacetonfosfat omdannes til 3-fosfoglycersyre via 3-fosfoglyceraldehyd som gir NADH og 1,3-difosfoglycersyre som igjen gir ATP og 3-fosfoglycersyre. På denne måten kommer ATP og NADH ut i cytplasma. 3-fosfoglycersyre fraktes inn i kloroplasten i bytte mot fosfat som igjen reduseres til dihydroksyacetonfosfat i kloroplasten.
Dikarboksylattranslokator
Dikarboksylattranslokator frakter dikarboksylsyrer som oksaleddiksyre, malat, aspartat og 2- oksoglutarat over kloroplastmembranen. Kan brukes til å frakte reduserende ekvivalenter over membranen
H+/ATP er 4. Q-syklus ekstra protoner. Fotosyntese fra 1 mmol m-2 s-1 for skygge planter opptil 50 mmol for solplanter.
Oppkonsentrering av CO2 i planten
Konsentrasjonen av CO2 i vann i likevekt med luft er ca. 10 mM. Sammenlignet med dette er Km- verdien for CO2 til rubisko mellom 15-25 mM hos C3-planter og hos blågrønnbakterier kan Km- verdien være opptil 200 mM. Derimot er PEP karboksylase mettet med substrat (HCO3-) under de samme betingelsene. Hos C4-plantene oppkonsentreres CO2 i slirecellene via transport av C4-syrer. I motsetning til hos planter hvor rubsiko er spredt rundt i stroma i kloroplastene ligger rubisko i karboksysomer hos blågrønnbakterier og i pyrenoid i alger.
Karbonsyre anhydrase er et zinkenzym som katalyserer likevekten mellom bikarbonat og karbondioksid:

Moroney, J.V. & Somanchi, A.: How do algae concentrate CO2 to increase the efficiency of photosynthetic carbon fixation ? Plant Physiol. 119 (1999) 9-16.
Lys gir gir også mer basisk stroma og økning i konsentrasjonen av Mg2+. Høyere pH passer bedre med pH optimum for enzymene i Calvin syklus.
Triosefosfat byttes mot fosfat via fosfattranslokatoren i kloroplastmembranen. Tilgangen på fosfat kan derved også være med å regulere Calvin-syklus.
Figurer
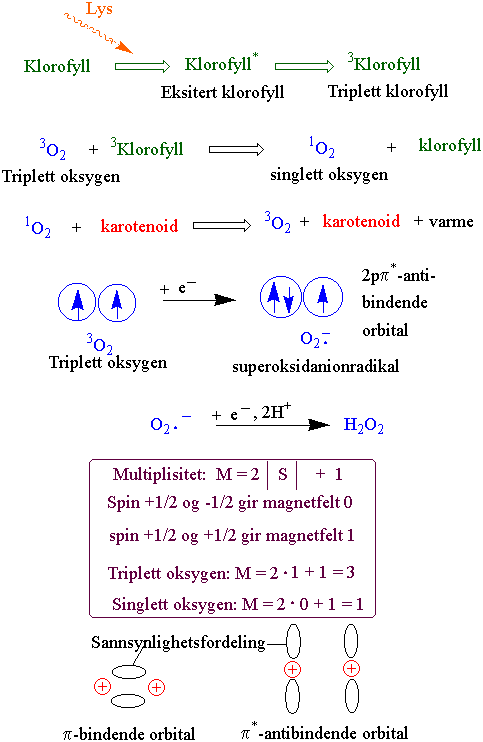{kind=link}
{kind=link}