Plantene kan overføre signaler fra omgivelsesfaktorene rundt plantene til en fysiologisk respons inne i plantene. Signalmolekyler deltar i en signaloverføringsvei.
Signaloverføring hos prokaryoter
Bakterier kan raskt tilpasse seg endringer i omgivelsene. Bevegelige bakterier kan flytte seg i en gradient av ytre omgivelsesfaktorer. For å kunne reagere på et ytre signal må:
- 1) Bakterien kunne registrere det ytre stimulus.
- 2) Stimulussignalet må forsterkes og overføres.
- 3) det må føre til en ønsket respons.
Bakterier har en signaloverføringssystem basert på to komponenter. Bakteriene registrerer kjemiske stoffer (ligander) i miljøet ved hjelp av receptorer på celleoverflaten. Ligander bindes til sensorproteiner i plasmamembranen eller i periplasmarommet mellom cellevegg og plasmamembran. Signalet fra det ytre stimulus overføres til et responsmottakerprotein ved at det skjer konformasjonsendring i sensorproteinet ved binding av liganden. Overføringen av signalet skjer ved fosforylering av et mottakerdomene og sensorproteinet har et transmitterdomene som virker som en autofosforylerende histidin kinase. Slike tokomponentsystemer har også blitt funnet hos gjær og planter. Fytokrom har blitt funnet i bakterier og responderer på rødt og mørkerødt lys og disse ligner på bakterielle sensorproteiner.
Signaloverføring hos eukaryoter
Dyrene utviklet et nervesystem og et endokrint system for å kunne tilpasse seg endringer i omgivelsene. Plantene utviklet ikke noe nervesystem, men et hormonsystem. For å kunne gi virkning må plantehormonene bindes til en reseptor (mottakermolekyl). Vanligvis er reseptorene proteiner festet til membraner. Steroidhormonene er hydrofobe og kan passere plasmamembranen (figur). Reseptorene som binder steroidhormoner virker som transkripsjonsfaktorer, DNA-bindende proteiner som er istand til å starte transkripsjon av bestemte gener etter at hormonet er bundet. Hormonpeptider kan virke ved å binde seg til plasmamembranen. Reseptorene for plantehormonene gibberelliner og abscisinsyre sitter celleoverflaten. For plantehormonet auxin finnes reseptorene både på celleoverflaten og i cytoplasma. Vanligvis overføres signaler i cellen indirekte via sekundære budbringere. De vanligste sekundære budbringere i plantene er 3´-5´ syklisk GMP (cGMP), inositol 1,4,5,-trifosfat (IP3) og 1,2- diacylglycerol (DAG) fra fosfolipider, syklisk ADP-ribose (cADPR) laget fra nikotinamid adenin diunkleotid (NAD+) og kalsium (Ca2+). Binding av hormoner til reseptorer på celleoverflatene gir en kjede av hendelser som leder til kortlevete sekundære budbringere (figur). Syklisk AMP (cAMP) spiller ikke samme viktige rolle i planter som i dyr.
{kind=link}
{kind=link}
{kind=link}
Reseptorproteinene reagerer med GTP-bindende regulatorproteiner kalt G-proteiner. Reseptorproteinet har G-proteinbindende domener. Både bakteriorhodopsin hos den fotosyntetiserende bakterien Halobacterium, rhodopsin i øyet hos vertebrater og luktreseptorer er eksempler på reseptorproteiner med 7 transmembranløkker med a-helix. G-proteiner består av 3 subenheter alfa, beta og gamma. I inaktiv form er GDP bundet til alfa-subenheten til G-proteinet og når G- proteinet bindes til reseptor-ligandkomplekset byttes GDP med GTP. Alfa-subenheten frigis fra G- proteinet og binder seg til adenylat syklase som aktiverer syntese av syklisk AMP (cAMP). Vanligvis er nivået av cAMP lavt i cellen katalysert av syklisk AMP fosfodisterase som hydrolyserer cAMP til 5´-AMP.
Konsentrasjonen av kalsium (Ca2+) er vanligvis lav i cellen, men ved åpning av Ca2+ kanaler kan konsentrasjonen plutselig øke.
Inositoltrifosfat er en budbringer som frigis fra plasmamembranen som respons på et hormonsignal og aktiverer protein kinase i cytoplasma. Hormoner og sekundære budbringere er en integert del av signalkjede og overføring fra ytre miljøbetingelser og respons på cellulært nivå. myo- inositol er et viktig mellomprodukt i plantenes metabolisme. Inositolheksafosfat er et viktig lager for fosfat i frø. I membranen finnes lipider som inneholder inositol. Galaktinol lages fra UDP- galaktose og inositol og er utgangspunktet for å lage raffinosesukker i platene.
{kind=link}
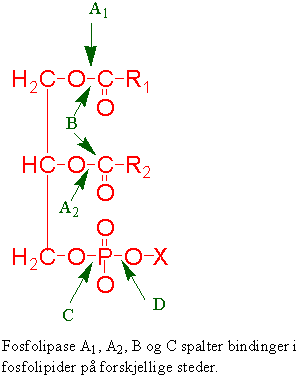
Fosfatidylinositol er et fosfolipid i plasmamembranen som kan fosforyleres to ganger ved hjelp av ATP katalysert av kinaser slik at det dannes fosfatidylinositol 4,5-bisfosfat. Når fosfatidylinoistol 4,5-bisfosfat spaltes av fosfolipase C dannes inositol 1,4,5-trifosfat (IP3) og diacylglycerol (DAG). DAG aktiverer protein kinase C og IP3 gjør at kalsiumkanalene i endoplasmatisk retikulum og vakuolemembran (tonoplast) åpnes. Prosessen slutter ved at IP3 brytes ned av en fosfatase. Kalsiumsignalet beveger seg som en oscillerende bølge gjennom cytosol.
{kind=link}
Noen proteinkinaser kan aktiveres av et kompleks av kalsium (Ca2+) bundet til proteinet kalmodulin. Konformasjonsendringen som skjer når Ca2+ bindes aktiverer Ca2+-kalmodulin avhengige protein kinaser som fosforylerer aminosyrene serin og threonin på enzymer og derved aktiveres enzymene.
Sekvenseringsdata fra Arabidopsis antyder at planter har en stor familie med reseptorlignende protein kinaser. Eksempler på slike er S-reseptor protein kinase som deltar i utskillelsen av glykoproteiner i forbindelse med selvinkompatibilitet i pollinering og befruktning hos planteslekten kål (Brassica).
Oligosakkarider, celleveggfragmenter virker også som elicitorerer. Bakterielle lipooligosakkarider er viktige signalmoleykler i induksjonen av nitrogenfikserende symbiose med erteplanter. Gener som koder for reseptorene for plantehormonene cytokinin og etylen inneholder domener med autofosforylerende histidin kinase.
Protein kinaser
Cellereseptorer overfører ekstracellulære signaler til en respons inne i cellene. Reseptorene som sitter på overflaten til cellene kan virke direkte som enzymer eller være koblet til andre enzymer. De vanligste signalreseptorene er:
1) Kjemisk styrte ionekanaler som åpner ionekanaler når et de binder et kjemisk stoff. Det er disse som åpner ionekanaler i overføring av nerveimpulser når de binder en neurotransmittor.
2) G-protein reseptorer er proteiner som kan binde og hydrolysere GTP, hvor omdanning av GTP til GDP vil gi en respons. Disse proteinene har en løkke som går 7 ganger gjennom membranen. De ble oppdaget av M. Rodbell og A. Gilman som fikk nobelprisen i medisin i 1994 for dette arbeidet. G-protein reseptorer kan bl.a. aktivere fosfolipase.
3) Protein kinaser er en stor familie med proteiner som overfører g-fosfatgruppen fra ATP til aminosyrene serin, threonin, tyrosin og histidin i proteiner. Aktiviteten og responsen oppheves og balanseres av spesifikke protein fosfataser som fjerner fosfatgruppen igjen. Calcineurin er en protein fosfatase som aktiveres av Ca2+-calmodulin og hemmes av cyklosporin og derved påvirker immunsystemet. Cyklosporin kan også påvirke kalsiumindusert inaktivering av K+-kanaler i lukkecellene i spalteåpningene hos planter.
14-3- 3- proteiner er proteiner som kan lette fosforyleringen. 14-3-3-proteiner er en proteinfamilie som binder seg til andre proteiner og deltar i signaloverføring.
Protein kinaser styrer store deler av metabolismen og påvirkes av en rekke omgivelsesfaktorer rundt planten. Virkningen av hormoner skjer ofte via protein kinaser. Det katalyttiske domene på protein kinasene består av en konservert sekvens på ca. 300 aminosyrer. Protein kinasene kan deles i grupper:
a) Reseptorlignende protein kinaser (RLK) som har 3 proteindomener: et ekstracellulært utenfor cytoplasma, en del som går igjennom membranen og en del inne i cytoplasma. Binding av en ligand til receptoren gir en dimerisering av proteinet. Protein kinasene kan også danne et proteinkompleks med en protein fosfatase.. Eksempler på RLK er:
- a1) S-domene RLK som har et ekstracellulært domene som ligner på glykoproteiner som lages av S-locus i kål (Brassica) og deltar i sporofytt inkompatibilitet.
- a2) Epidermal vekstfaktorlignende RLK (EGF-RLK) som bl.a. finnes i cellevegger.
- a3) Leucinrike repeterte RLK (LRR-RLK) som inneholder en kjerne med mye av aminosyrene leucin og asparagin.
- a4) Lektin reseptor kinase RLK (LR-RLK) som bl.a. binder oligosakkarider og lektiner (glykoproteiner).
b) Kalmodulin kinase avhengige protein kinaser (CaMK) som trenger kalsium bundet til proteinet kalmodulin. Disse finnes bl.a. i cytoplasma, cytoskjelett og plasmamembranen. Eksempler er CpCK1 som kan gi myristylering (binding av mysistinsyre). Sukrose ikkefermenterende kinaser (SNF) som styrer forholdet mellom anabolisme og katabolisme.
c) CMGC- gruppen som styrer vekst og utvikling.Omfatter mitogenaktiverte protein kinaser (MAPK) som påvirker transkripsjonsfaktorer og deltar i en kaskadereaksjon med protein kinaser. Kalmodulin kinase II (CKII) som er funnet i hjernen og også i planter regulerer den transkripsjonen. Ofte er det slik at når transkripsjonsfaktorene er fosforylert kan de binde seg til DNA. MAPK deltar i overføring av signaler fra stress, for eksempel mekanisk berøring, osmotisk stress og såring. Andre eksempler i denne gruppen er syklin-avhengig protein kinase (CDK) som deltar i cellesyklus, samt glykogen syntase protein kinaser (GSK). En signaloverføringsvei kan være som følger signal →Ras → MAPKKK (raf) → MAPKK → MAPK → transkripsjon
d) Raf med bl.a. CTR1 (konstitutiv trippel respons) som er blitt oppdaget i etyelnufølsomme mutaner.
e) AGC-gruppen med eksempel PVK1