Hos vannlevende dyr blir ammonium utskilt og fortynnet i vannmassene, men landlevende dyr skiller ut nitrogen i form av urea eller urinsyre for å unngå giftvirkningen av ammonium. Ammonium blir produsert ved katabolisme av protein og aminosyrer.
Ammoniumtoksisitet og planter
Ammoniumfølsomme (ammoniumsensitive) planter får morfologiske endringer og redusert vekst av skudd og røtter i nærvær av høye konsentrasjoner av ammonium, til forskjell fra ammoniumtolerante planter. Planter som vokser på sur jord i barskog, slik som bartrær og lyngplanter er ammoniumspesialister som tar opp nitrogen fra jorda i form av ammonium. Grunnen til at ammonifiseringen stopper ved ammonium er liten nitrifikasjon grunnet sur humus og silikatrik podsoljord med podsolprofil med liten bufferkapasitet, som gir dårlige vekstforhold for nitrifiserende bakterier. Nedbrytningen av organiske nitrogenforbindelser skjer vesentlig via sopp som er mer tolerante for lav pH, men ammonium blir ikke vider oksidert til nitrat. Opptaket av ammonium gjør at jorden blir surere, i tillegg til forsuringen fra nedbrytningen av surt strøfall. Ved gjødsling av skog bruker man derfor urea som nitrogenkilde. Fra hagen er rhododendron et eksempel på en surjordsplante som ikke skal ha nitratgjødsel, men ammoniumgjødsel.
Generelt kan bruk av urea, ammonium og ammoniakk som N-kilder gi uheldige effekter hos planter som ikke har ammoniumtoleranse.
Et annet økosystem med lav nitrifikasjon er anaerob jord og anaerobe sedimenter uten oksygen, for eksempel i oversvømte rismarker. Nitrifikasjon er avhengig av oksygen. Det betyr at nitrogenkilden for våtlandsris blir ammonium, og derfor brukes også her urea som mineralgjødsel. I jorden er det ekstracellulære enzymer som hydrolyserer urea ((NH2)2CO), men reaksjonen kan også skje ikke-enzymatisk :
\((NH_2)_2CO \;+\; H_2O \rightarrow \; 2\;NH_3 + \; CO_ 2\)
Det er indikasjoner på at ammoniumspesialister som gran kan akkumulere og lagre større ammonium i sure kompartementer i cellene, studier gjort med N-14 og N-15 NMR. Ris (Oryza sativa) er eksempel på en art som tåler å vokse i nærvær av høye konsentrasjoner med ammonium, men bygg (Hordum vulgare), erter (Pisum sativum) og andemat (Lemna sp.) mistrives med ammonium som eneste N-kilde.
Ammoniakk (NH3) finnes både som gass og væske. Ammoniakk er meget flyktig og fordamper lett, i motsetning til ammonium som er i ioneform.
NH3 + H2O ⇔ NH4+ + OH-
Den basiske ionisasjonskonstanten Kb:
\(K_b= \displaystyle\frac{ [NH_4^+] [OH^-]}{[NH_3]}\)
\(pOH = pK_b + \log_{10}\displaystyle\frac{\text{konjugert kation}}{\text{base}}= pK_b - \log_{10}\displaystyle\frac{\text{base}}{\text{konjugert kation}}\)
Væske det vil si vann (H2O) inngår ikke i likevektsuttrykket
NH4+ + H2O ⇔ NH3 + H+
\(K_a= \displaystyle\frac{ [NH_3] [H^+]}{[NH_4^+]}= 5.6\cdot 10^{-10}\)
Den sure ionisasjonskonstenten eller likevektskonstanten Ka
\(pK_a= -\log_{10}K_a= \log_{10}5.6\cdot 10^{-10}= 9.25\)
Henderson-Hasselbach ligningen
\(pH= pK_a +\log_ {10} \displaystyle\frac{\text{konjugert base}}{\text{syre}}\)
Når det er like mengder konjugert base (NH3) som syre (NH4+) så er pH lik pKa-verdien som vist på Bjerrumdiagrammet nedenfor. Log10(1)= 0
Nitratplanter
Nitratplanter (nitrofile planter) som foretrekker nitrat som N-kilde, og som vokser på nærings- og moldrik jord med nitrifiserende bakterier, er dem som oftest ikke tåler ammonium som N-kilde. Imidlertid er det slik at de fleste planter kan bruke både ntirat og ammonium som uorganiske nitrogenkilder, og spesielt hvis det bare er lave konsentrasjoner av ammonium. Når plantene har tatt opp nitrat blir det redusert til ammonium i en nitratreduksjon. I plantene er det fler enzymsystemer som deltar i fikseringen av ammonium, slik at det ikke foreligger fritt ammonium i cellene. Ved fotorespirasjon blir det produsert ammonium, men også denne blir reassimilert. Nitratplanter kan lagre større mengder nitrat i vakuoler, men planter har ikke samme evne til å lagre ammonium, bortsett fra hos gran (Picea abies) hvor vi har funnet indikasjoner på at dette skjer. Det finnes både høyaffinitets- og lavaffinitetsopptakssystemer for nitrat, tilpasset henholdsvis lave eller høye konsentrasjoner av nitrat. På samme måte finnes det høyaffinitets- og lavaffinitetsopptakssystemer for ammonium hos ammoniumspesialistene.
Hvorfor er ammonium giftig ?
En av forklaringene på hvorfor ammonium (NH4+) er giftig for planter som ikke er adaptert til å vokse på sur jord er at opptak av ammonium gir forsuring. Forsuringen ved ammoniumopptaket kan være betydelig hvis det er dårlig bufferkapasitet i jorden.

Opptak av ammonium gir forsuring (NH4+ - H+ antiport), mens opptak av nitrat gir høyere pH (NO3- - H+ symport).
Sur jord gir økt tilgang på jern, og det er en også hypotese om at for sur jord gir for mye jern slik at giftvirkningen ikke direkte er via pH, men skyldes for mye jern. Alle organismer i en oksygenatmosfære har vanskeligheter med å ta opp jern, og planter som vokser i mer basisk jord har et velutiklet opptakssystem for jern, og det er mulig at dette systemet blir overbelastet hvis pH synker.
Den andre forklaringen er pH-likevekten mellom ammonium (NH4+) og ammoniakk (NH3).
\(NH_4 ^+ \;\; \iff\;\; NH_3 \;+\; H^+ \;\;\;\;\;\, pK_a = 9.25\;\;\;\; (20^oC)\)
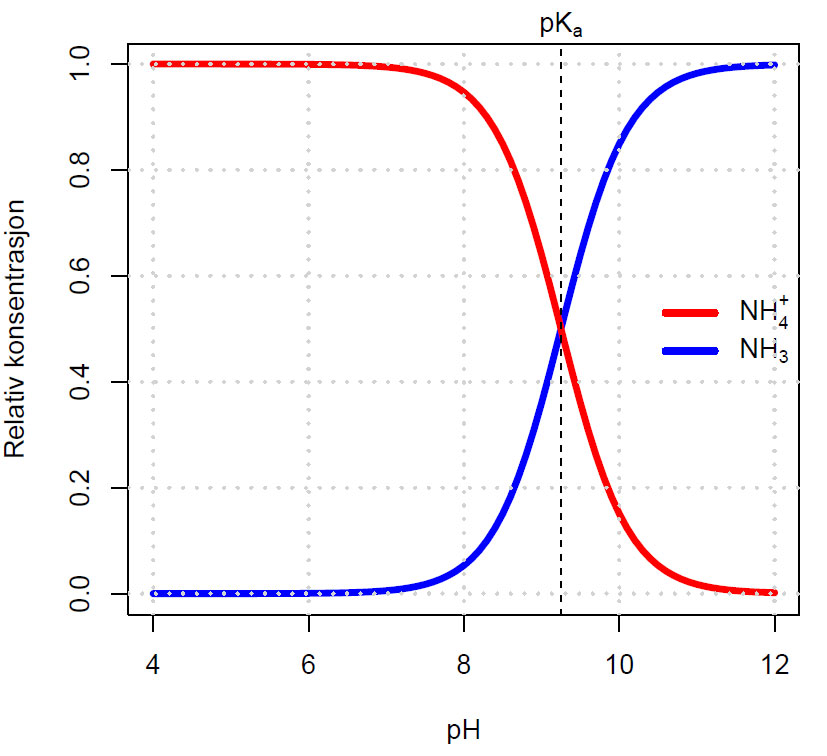
Relativ konsentrasjon av de to nitrogenformene ammonium (NH4+) og flyktig ammoniakk (NH3) som funksjon av pH (Bjerrumdiagram).
Det betyr at når pH i cellene er høyere enn 7 så begynner noe av nitrogenet å foreligge i form av ammoniakk (NH3) som er et nøytralt molekyl, og som kan diffundere lett gjennom membraner og forstyrre H+-gradienter over membraner og membranpotensialet Ammoniakk har stikkende lukt som advarsel om giftighet, og man kan registrere 5 mM konsentrasjoner ammoniakk i luften , jfr. utslipp fra kjøleanlegg basert på ammoniakk, samt salmiakk som rengjøringsmiddel. I husdyrgjødsel som har høy pH så vil mye av nitrogenet fordampe ut i atmosfæren som ammoniakk. Ammoniakk har også vært brukt til ensilering av gras til husdyrfor.
Ammoniumforgfitning hos dyr
Ammonium som blir produsert i katabolismen av aminosyrer, puriner og pyrimidiner må bli fjernet for å unngå skade. Dyr som lever akvatisk i saltvann (marint) eller ferskvann (limnisk) skiller ut ammonium ut i vannmassene hvor det blir fortynnet (ammonoteliske dyr). Terrestriske pattedyr skiller ut ammonium som urea (ureoteliske dyr), mens fugler og krypdyr (reptiler) skiller ut nitrogen i form av urinsyre.Ammonium og ammoniakk er giftig for alle virveldyr (vertebrater) og påvirker nervessystemet ved depolarisering av membranpotensialet i nevroner via endringer av transport av kalium (K+) og kalsium (Ca2+) gjennom spennings- og ligandregulerte ionekanaler, samt påvirker nevrotransmittorer som NMDA glutamatreseptorer. Dyr i vann kan kvitte seg med ammonium ved å skille det ut i vannet, men fisk som lever tett sammen i oppdrettsanlegg eller akvarier kan bli utsatt for giftige konsentrasjoner med ammonium. En annen strategi er å omdanne ammonium til urea og/eller urinsyre.
Aminosyren glutatmat kan omdannes til glutamin i en glutamat-glutaminsyklus i astrocytter (stjerneformete gliceller) og neuroner hjernen.
Alle organismer inneholder enzymet glutamin syntetase som katalyserer en reaksjon mellom aminosyren glutamat og ammonium hvor aminosyren glutamin blir dannet:
glutatmat + NH4+ + ATP → glutamin + ADP + Pi + H+
Dette er en viktig enzym i assimilasjonen av ammonium i bakterier og planter, men hos pattedyr deltar det i avgiftningen av ammonium hvor glutamin kan fraktes via blodet til videre omsetning i lever og nyrer.
Glutamat og gamma-aminosmørsyre (GABA) er viktige nevrotransmittorer i hjernen, og hører til glutamatfamilien av aminosyrer hvor ammonium er en av metabolttene som må holdes under kontroll. Det betyr at det må opprettholdes en ammoniumlikevekt (ammoniahomeostase). Ammoniakk (NH3) kan diffundere lett over lipidmembranene i hjernen. Aminosyretransportører, og en rekke forskjellige enzymer deltar i omsetningen (e.g. transaminaser, glutamat dehydrogenase).
Ammoniumforgiftning hos mennesker
Det er lave konsentrasjoner av ammonium i blod (< 50 µM), men 0.1 mM NH4+ kan føre til forgiftning og bevisstløshet (hjernen er meget følsom for ammonium). Giftvirkningen av ammonium og ammoniakk (NH3) kan muligens skyldes interferering med trasport av kalium (K+) som blir forstyrret når ammoniakk forstyrrer pH-gradienter og membranpotensial. Ammonium fra metabolismen blir omsatt i leveren til urea som blir utskilt med urinen.
Kilder til ammonium hos dyr og mennesker
Katabolisme av L-aminosyrer kan gi opphav til ammonium
Glutaminase
Ammonium blir laget ved katabolisme av aminosyren glutamin i lever og nyrer katalysert av enzymet glutaminase som omsetter aminosyren L-glutamin til aminosyren L-glutamat (glutaminsyre) og ammonium (NH4+):
glutamin + H2O → glutamat + NH4+
Enzymet finnes som tre forskjellige isoenzymer lokalisert til til forskjellige kroppsorganer og i mitokondirer. Enzymet kan regulere mengden av nevrotransmittoren glutamat i hjernen, samt deltar som en del av systemet for kontroll av pH (pH-homeostase).
Glutamat dehydrogenase
Glutamat dehydrogenase ktalyserer reaksjonen:
glutamat + NAD(P)+ ↔ NH4+ + α-ketoglutarat (2-oksoglutarat) + NAD(P)H + H+
Enzymet glutamat dehydrogenase finnes hos både prokaryoter og eukaryoter. Enzymreaksjonen er reversibel og kan koble sammen anabolisme og katabolisme av aminosyrer. Hos dyr kan ammonium bli assimilert via glutamat dehydrogenase kombinert med enzymet glutamin syntetase. Hos dyr er ADP-ribosylering en del av reguleringsmekanismen av enzymet. Alfa-ketoglutarat kan inngå i transamineringsreaksjoner, men kan også omsettes i sitronsyresyklus for å skaffe energi. Det er eksempel på anaplerotisk reaksjoner hvor et mellomprodukt (intermediat) inngår i flere omsetningsveier i metabolismen. Glutamat dehydrogenase har en høy Km-verdi for ammonium, slik at man antar at enzymet deltar i katabolisme av glutamat i stedet for den reverse reaksjonen med assimilasjon av ammonium.
Ketosyrene kan inngå i transamineringsreaksjoner:
aminosyre (aminodonor) + 2-ketosyre (aminoakseptor) ↔ produkt ketosyre + produkt aminosyre
Alle aminosyrer bortsett fra lysin kan transamineres
Aminosyre oksidase
Aminosyre oksidase er enzymer som oksiderer aminosyrer hvor ammonium, en oksosyre (ketosyre) og hydrogenperoksid (H2O2) er produkter i reaksjonen, en oksidativ deaminering av L-aminosyrer. Det er flavoenzymer med FAD-bindende sete.
L- aminosyre + O2 + H2O → NH4+ (NH3) + 2-oksosyre (α-ketosyre) + H2O2
Aminosyre oksidase er også et enzym som man kan finne i slangegift (Viperidae, Elapidae, Crotalidae)
Alanin oksidase
L-alanin + O2 + H2O → NH4+ + pyruvat + H2O2
Glycin oksidase:
glycin + O2 → NH4+ + glyoksylat + H2O2
Serin deaminase
Serin deaminase (L-serin ammonialyase) er et pyridoksalfosfatenzymet som omsetter L-serin til pyruvat og ammonium.
serin → NH4+ + pyruvat
Nedbrytning av nukleinsyrer
Puriner
Adenosin deaminase og guanin deaminase er enzymer som deltar i metabolismen av puriner, og i nedbrytning av henholdsvis adenosin og guanosin i maten. Enzymet inneholder zink og har også funksjoner i immunsystemet.
Adenosinmonofosfat blir omdannet adenosin katalysert 5\-nukleotidase
adenosin monofosfat + H20 → adenosin + Pi
Enzymet aeenosin deaminase omdanner adenosin til inosin.
adenosin + H2O → inosin + NH4+ (NH3)
Enzymet guanosin deaminase katalyserer reaksjonen
guanosin + H2O→ xanthin + NH4+(NH3)
Ribose fjernes fra inosin katalysert a ven nukleosidase
Inosin + H2O ↔ hypoxanthin + ribose
Xanthin oksidase katalyserer deretter trinnene:
hypoxanthin + H2O + O2 ↔ xanthin + H2O2
xanthin + H2O + O2 ↔ urinsyre + H2O2
Fugler, krypdyr, insekter og pattedyr kan skille ut urinsyre. Xantin er en forløper og mellomprodukt i biosynteseveien av urinsyre, Urinsyre er kjemisk likt bygget opp som puriner, en av nukleinsyrene . Urinsyre inneholder fire nitrogenatomer og kan danne krystaller. Xantin oksidase er enzymet som lager urinsyre fra xantin og hypoxantin. Enzymet inneholder molybden. Guano er ekskrementer fra fugler og huleboende flaggermus brukt som nitrogen og fosfor-gjødsel i landbruket.
Urinsyre kan bli videre oksidert til allantoin katalysert av urat oksidase lokalisert til peroksisomer:
urinsyre + O2 + H2O → allantoin + CO2
Urat oksidase finnes fra bakterier til sopp, planter og pattedyr, men både menneskeaper og mennesker mangler dette enzymet.
Allantoin blir videre omdannet til allantoat katalysert av allantoinase.
Allantoikase omdanner allantoat til urea
Allantoat + H2O → urea + glyoksylat
Amfibier og bruskfisk kan skille ut urea laget på denne måte, altså ikke via ureasyklus.
Vannlevende organismer kan skille ut ammonium i vannmassene laget fra urea katalysert av enzymet urease
2urea + 2H2O → 4NH4+ + 2CO2
Ved nedbrytning (katabolisme) av makromolekyler som inneholder nitrogen så lagres og benyttes karbon, hydrogen, oksygen, men overskudd av nitrogen skilles ut. Ammonium kommer fra nedbrytning av proteiner, spesielt deaminering av aminosyrer. Ammonium kan gi en rekke fysiologiske og skadelige effekter på muskler og nervesystem i kroppenL anddyr må avgifte ammonium ved å omdanne det til mindre giftig vannløselige urea og urinsyre, henholdsvis ureoteliske og urikoteliske dyr. Evolusjonære forskjeller mellom invertebrater, pattedyr, fugler og krypdyr hvordan de kvitter seg med overskudd av nitrogen.
Puriner
Ved nedbrytning av pyrimidiner e.g. thymin blir det i reaksjonen fra β-ureidoisobutyrat produsert ammonium katalysert av β-ureidopropionase, hvor β-alanin også kan være et substrat
β-ureidoisobutyrat + H2O → β-aminoisobutyrat + NH4+ + HCO3-
Tilsvarende for nedbrytning av cytosin.
Assimilering av ammonium
Ammonium frigitt i biokjemiske reaksjoner må raskt bli reassimilert for å unngå toksiske reaksjoner. Reassimileringen er en del av anabolismen også en del av katabolismen og sekresjonsprosjektet hvor ammonium, urea eller urinsyre blir skilt ut fra kroppen ved ekskresjon eller avføring.
Glutamat dehydrogenase katalyserer oksidativ deaminering av glutamat til ammonium og alfa-ketoglutarat. I revers bruker alfa-ketoglutarat fra sitronsyresyklus og reduserr syntesen av ATP
Glutamat dehydrogenase
Alfa-ketoglutarat + NH4+ + NAD(P)H ↔ glutamat + NAD(P)+
Glutamin syntetase
Glutamin syntetase (GS) katalyserer en ATP-avhengig assimilasjon av ammonium inn i aminosyren glutamat som blir omdannet til glutamin.
Glutatmat + NH4+ + ATP → glutamin + ADP + Pi (H2PO4-)
Glutamin syntetase finnes i alle levende organismer, både prokaryoter og eukaryoter og i form av flere isoenzymer. I planter deltar det i nitrogenassimilasjonen. Ugrasmiddelet fosfinotricin (glufosinat) hemmer GS ved å binde seg til sete på enzymet som binder glutamat. Metionin sulfoksimin (MSO) er et annet eksempel på en inhibitor som binder seg til glutamatbindingssete. MSO kan deretter bli fosforylert med ATP og lage irreversibel hemming. Hos dyr finnes glutamin syntetase i lever, nyrer og hjerne hvor det deltar i avgiftning av ammonium. Astrocyttene i hjernen beskytter nevronene mot ammoniumtoksisitet ved å ta opp ammonium og glutamat.
I hjernen kan GS delta i resirkulering av nevrotransmittoren glutamat. Glutamin kan bli brukt biosyntesen av nukleinsyrer (puriner) .Lever har en viktig rolle i nitrogenmetabolismen og avgiftningen av ammonium. Dette enzymet spiller en viktig rolle i fjerning av ammonium i andre deler av kroppen lokalt
I oppdrett, husdyrbruk og muskeloppbygging kan det i tillegg til naturlige proteinkilder være tilsatt essensielle aminosyre bl.a. lysin og metionin. Det første trinnet i ureasyklus er karbamoylfosfat syntase 1 i mitokondriene som har relativt lav affinitet for ammonium (Km-verdi1-2 mM) , men glutamin syntetase har høy affinitet for ammonium (Km-verdi 0.2 mM).
Aminosyren glutamin er en viktig intern transportform for nitrogen fra muskler og lever
Ved leverskader kan det være viktig å redusere inntaket av protein i maten.
Ammonium utskilt via urea i ureasyklus hos dyr
Ureasyklus (Hans Krebs og Kurt Henseleit-syklus) består av fem enzymatiske trinn aminosyren ornitin omdannes til forskjellige intermediater, og endeproduktet urea kan skilles ut og derved fjernes potensielt gifitig ammonium. Planter benytter bare enkelte av enzymtrinnene i ureasyklus til sine vekstformål
2NH3 + CO2 + 3ATP + H2O → H2N-CO-NH2 (urea) + 2ADP + AMP + 2Pi + PPi
Ureasyklus begynner i mitokondriene hvor hydrogenkarbonat (HCO3-) og ammonium danner kabamylfosfat ved bruk av 2 ATP.
Enzymet karbamylfosfat syntase katalyserer reaksjonen med assimilering av ammonium:
HCO3- + 2 ATP + NH4+ → karbamylfosfat
Finnes som to isoenzymer. Isoenzym 1 i mitokondriene som bruker ammonium fra proteinrik mat. Isoenzym 2 i cytosol som bruker glutamin som N-kilde og inngår i biosyntese av pyrimidiner, og hemmes av pyrimidinnukleotider.
I neste trinn reagerer kabamylfosfat med ikke-proteinaminosyren ornitin og danner citrullin atalysert av ornitin karbamyltransferase
karbamylfosfat + ornitin → citrullin
Citrullin kobles deretter sammen med aminosyren aspartat og danner arininosuccinat med ATP som energikilde og som degraderes til AMP og pyrofosfat (PPi), katalysert av argininsuccinat syntetase
Citrullin + aspartat + ATP → argininosuccinat + AMP + PPi
Enzymet argininsuccinase katalyserer reaksjonen for fumarsyre (fumarate) inngår og man får dannet den nitrogenrike aminosyren arginin:
argininosuccinat + fumarate → arginine
Enzymet arginase omdanner aminosyren arginin til urea og ornitin. Urea skilles ut med urinen og ornitin kan brukes på nytt i syklusen:
arginin + H2O → urea + ornitin
Urea (CO(NH2)2 er vannløselig. Dannelsen av urea krever kjemisk energi (ATP) og tilgang på vann for å fortynne urea og skille det ut hos ureotelisk dyr.
Genmodifiserte planter og ammonium
Aminosyren glutamat (glutaminsyre) spiller en essensiell rolle ved assimilasjonen av ammonium i planter. Det bredspektrede ugrasmiddelet glufosinat (phosphinothricin) hemmer ammoniumassimilasjonen hos både enfrøbladete og tofrøbladete planter. Glufosinat som selges under handelsnavnet Basta® er en naturlig aminosyre med en fosforgruppe, finnes i aktinobakterier av slekten Streptomyces, blant annet i tripeptidet bialaphos, Glufosinat (phosphinothricin) hemmer enzymet glutamin syntetase i plantene. Blokkering av glutamin syntetase gir opphopning av ammonium i plantevevet som gir giftvirkning, samt mangel på de andre aminosyrene dannet fra glutamat og glutamin, som resulterer at planten dør. Ved å sette inn genet for phosphinothricin acetyltransferase, isolert fra aktinobakteriern Streptomyces viridochromogenes , som lager proteinet som inaktiverer phosphinotricin, gjorde Bayer det mulig å lage genmodifiserte planter som tåler sprøytemiddelet, og derved kan opprettholde ammoniumassimilasjonen.