{kind=link}
Innen medisinsk behandling brukes begrepene bredspektret antibiotika om stoffer som stopper veksten av mange forskjellige arter bakterier ved å hemme sentrale fundamental metabolske funksjoner felles for store grupper bakterier, for eksempel Gram-positive bakterier eller Gram-negative bakterier.
Smalspektret antibiotika hemmer spesifikt bare noen få bakteriegrupper ved å interferer eller hemme noen få spesifikke enzymer eller reaksjoner som er spesifikk for bare noen få bakterieslekter.
Antivirale stoffer er moekyler som hindrer virus i å feste seg på utsiden av vertsceller ved å blokkere overflatereseptorer, ved å hindre genomreplikasjon og formeringen av virus når de har kommet inn i celler, eller ved å blokkere lysis av cellene som frigir virus fra cellene. Antivirale stoffer er ikke antibiotika, siden definisjonen av liv er knyttet til cellen
Antibiotikaresistens
Antibiotika påvirker metabolismeprosesser i bakteriene. Antibiotika kan lages mer effektive i den farmasøytiske industrien ved kjemisk å endre sidegrupper til molekylene slik at man får semisyntetiske antibiotika. Et antibiotikum som dreper både Gram-positive bakterier og Gram-negative bakterier kalles bredspektret antibiotika. Smalspektret antibiotika virker bare på noen få arter med bakterier. Fremtiden så lys ut, men mørke skyer skulle seinere dukke opp i horisonten. Det tok ikke lang tid fra den første bruk av antibiotika til det dukket om resistente bakteriestammer. Resistente bakterier som er tolerante mot antibiotika og hvor veksten ikke påvirkes. Dette er et naturlig resultat av darwinistisk (biologisk) evolusjon hvor antibiotika virker som et seleksjonspress. Med det enorme antall bakterier som finnes og den eksponentielle veksthastigheten tar det ikke lang tid før seleksjonsmekanismen og naturlig utvalg velger ut resistente (tolerante) bakterier. Antibiotika virker som et selektivt press, og darwinistisk evolusjon gir seleksjon av bakterier som har molekylære mekanismer eller enzymer som gjør dem resistente og tolerante for antibiotika. Gener for antibiotikaresistens kan være lokalisert til plasmider i bakteriene, og kan bli overført til andre bakterier via horistonal genoverføring. Antibiotikaresistens er godt eksempel på hvordan evolusjonen virker, for dem som tviler på evolusjon som mekanisme for seleksjon og artsdannelse. Utvikling av antibiotikaresistens skaper en betydelig utfordring for helsemyndighetene. Den farmasøytiske industrien kan lage kjemiske modifiseringer på naturlig forkommende antibiotika, eller lete nye typer antibiotika, men det er ikke et prioritert område. Det finnes mer enn 150 forskjellige antibiotika fordelt på mer enn 17 forskjellige hovedtyper. Noen tenker seg bruk av bakterievirus (bakteriofager) til å drepe bakterier, men det er mange praktiske vanskeligheter med gjennomføring.
Eksempler på antibiotikaresistente bakterier ermethicillin-resistente gule stafylokokker (Staphylococcus aureus, MRSA). Vancomycin-resistente Enterococcus (VRE). Multiresistente tuberkulosebakterier (Mycobacterium tuberculosis MDR-TB). Kcarbapenemase-produserende Enterobacterales (CPE) er resistente mot de fleste typer antibiotika. Karbapenemaser er β-lactamaser, enzymer med bred ved substratspesifisitet som kan hydrolysere mange typer antibiotika som penicilliner, cefalosporiner, monobaktamer og ogkarbapenemer. Gener som koder for karbapenemaser finnes blant annet på plasmider i bakterien Klebsiella pneumoniae.
Beta-laktam antibiotika
Penicilliner, cephalosporiner og cephamyciner er beta-lactam antibiotika inneholder en beta-laktam-ring og hemmer enzymene transpeptidaser i Gram-positive bakterier (Gramfarging) og derved blokkerer reaksjoner som har med kryssbinding av peptidoglykaner i celleveggen til bakteriene.

Aminoglykosidantibiotika
Aminoglykosidantibiotika inneholder aminosukker bundet med glykosidbinding til andre aminosukker og gir veksthemming hos bakterier ved å feste seg til ribosomene og ødelegger translasjonen. Aminoglykosidantibiotika hemmer også proteinsyntese i mitokondrier og kloroplaster, organeller som er prokaryot opprinnelse. Omfatter kanamycin, gentamicin, neomycin og streptomycin.
Makrolidantibiotika
Macrolidantibiotika inneholder store laktonringer bundet til sukker. Omfatter erythromycin, oleandomycin, spiramycin og tylosin.
Tetrasykliner
Tetracykliner som er laget av et naftalenringsystem (klortetracyklin, oksytetracyclin) hemmer vekst av både Gram-positive bakterier og Gram-negative bakterier og er således bredspektret antibiotika. Tetracyklin hemmer binding av aminoacyl-tRNA til ribosomene. Tetracyklin binder seg til 30S subenheten på ribosomene og blokkerer bindingen av aminoacyl-tRNA, og derved blokkeres proteinsyntesen. I 1948 isolerte plantefysiologen Benjamin M Duggar ved Universitetet i Wisconsin er stoff fra en aktinobakterie i jord, og ble kalt aureomycin på grunn av den gule fargen. Aureomycin har antibakterielle egenskaper. Seinere ble det funnet et lignende stoff, terramycin (5-hydroksytetrasyklin) fra aktinobakterien Streptomyces rimosus. Ved hydrogenolyse av aureomycin ble produktet tetrasyklin, og som seinere ble også isolert fra Streptomyces albo-niger. Seinere ble det laget mange forskjellige tetrasykliner ved å endre kjemisk sidegruppene på tetrasyklin.
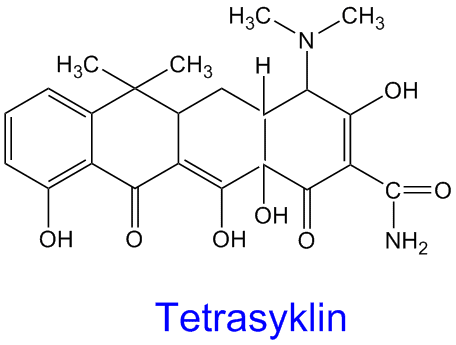
Kloramfenikol
Kloramfenikol er et bredspektret antibiotika som hemmer proteinsyntese ved å feste seg til 70S prokaryot ribosom og hemmer peptidyltransferase og hindrer derved at det dannes peptidbindinger. Kloramfenikol kan krysse blod-hjernebarriæren f.eks. i behandling av meningitis. Imidlertid har kloramfenikol som andre legemidler og antibiotika mange uheldige bivirkninger.
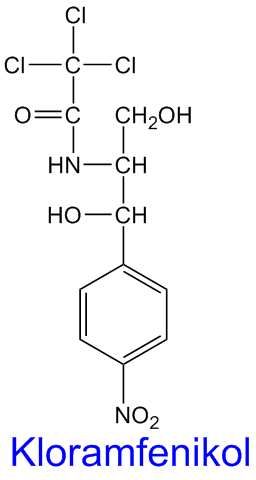
Sykloheksimid
Cykloheximid hemmer peptidyltransferase eukaryote ribosomer, og derved blokkeres eukaryot proteinsyntese. Sykloheksimid ble isolert fra aktinobakterien Streptomyces griseus. Imidlertid har sykloheksimid en rekke alvorlige giftige og teratogene bivirkninger og anvendes derfor ikke som legemiddel, men blir brukt i laboratorievirksomhet. Sykloheksimid gir DNA-skader og stoffet er en nyttig påminnelse om de mange giftige stoffene man kan finne i organismer, inkludert antibiotika.
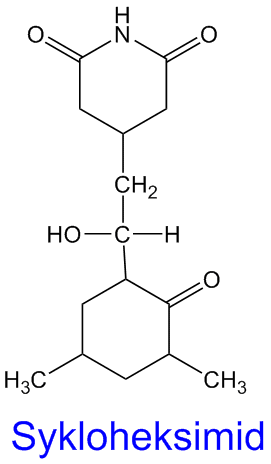
Streptomycin
Streptomycin er et aminoglukosid antibiotika som deltar i interaksjon med 70S ribosomer som gir blokkering av gjenkjennelse av mRNA-sekvensen og hemmer derved starten av proteinsyntesen (translasjonen). Ble under andre verdenskrig isolert fra aktinobakterien Streptomyces griseus. Et av de første antibiotika brukt i behandling av tuberkulose, nå gitt sammen med rifampicin, isoniazid og pyrazinamid, og streptomycin anvendes i dag mot en rekke bakterieinfeksjoner, for eksempel tularemi (Francisella tularensis), brucellose (Brucella sp.), Burkholderia infekesjon, samt pestbakterien Yersinia pestis) . Molekylet inneholder en inositol med to guanido-grupper festet til en metyl-pentose og videre til N-metyl-glukosamin
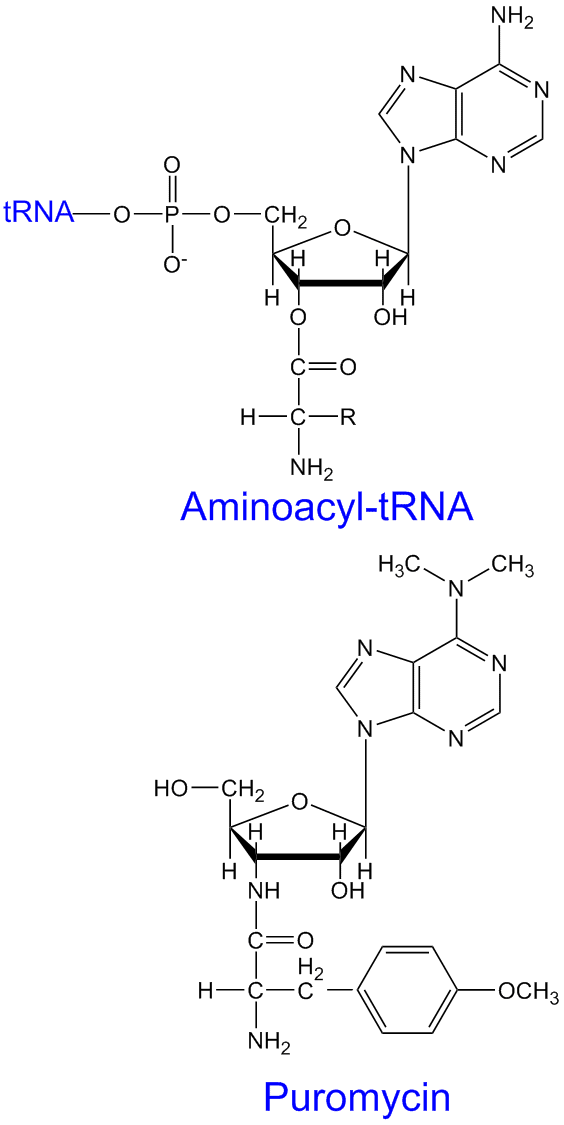
Puromycin
Puromycin virker som en analog til aminoacyl-tRNA på A-sete som hindrer dannelse av peptidbindinger og gir derved stopp av proteinsyntesen både hos prokaryoter og eukaryoter.
Gramicidin er en gruppe av lineære polypeptidantibiotika. Virker på Gram-positive bakterier ved å øke permeabiliteten av ioner over cellemembranen. Hører med til gruppen antimikrobielle peptider.
Azaserin er et antibiotika som hemmer purinbiosyntese og fører til kromosombrudd.
Actinomycin D er et antibiotika som binder seg til DNA og hindrer dannelse av mRNA.
Vancomycin påvirker dannelsen av cellevegg hos bakteriene ved å binde seg til peptidoglukan som gir veggen svakere struktur.
Oppdagelsen av antibiotika
Den britiske legen og bakteriologen Alexander Fleming oppdaget på 1920-tallet at en art muggsopp i slekten Penicillium kunne drepe en kultur med bakterier. Når muggsoppen vokste på agarskåler sammen med bakterier, ble det en sone rundt muggsoppen hvor bakteriene ikke kunne vokse. Et eksempel på den naturlige krigføringen mellom organismegrupper.

Professor Howard Florey ved Universitetet i Oxford startet i 1938 dyrking av muggsopp og isolering av penicillin, det antibakterielle stoffet Fleming hadde oppdaget i Penicillium notatum. Sammen med Ernst Boris Chain og medarbeidere ble de istand til å finne den kjemiske strukturen og igangsette kliniske studier.
Man var bekymret for giftvirkningen av penicillin, og i 1941 startet intravenøs behandling med penicillin av en svært syk pasient med sår i munnen som var blitt infisert med stafylokokker (Staphylococcus) og streptokokker (Streptococcus). Pasienten ble raskt bedre, en mirakelmedisin. Andre verdenskrig var i gang og det ble stort behov for store mengder antibiotika. Utover på 1940-tallet ble andre antibiotika isolert som streptomycin, kloramfenikol og cephlosporin. Fleming, Florey og Chain mottok i 1945 nobelprisen i medisin og fysiologi for oppdagelsen av antibiotikas helbredende virkning mot bakterieinfeksjoner. Mennesket hadde fått et verktøy til å bekjempe en lang rekke fryktede bakteriesykdommer, bl.a. lungebetennelse, bakteriell hjernehinnebetennelse blodforgiftning (sepsis), samt infeksjoner i bein og ledd. Sepsis er en alvorlig svikt i indre organer og deres virkemåte som resultat av en infeksjon. Folketallet i vesten får en ny oppsving som resultat av redusert dødelighet. Antibiotika som colistin er et polymyksin som man allerede tidlig oppdaget var giftig for nyrene. Noen antibiotika trenger ikke inn i cellevevet i kroppen og gjør at bakterier kan leve beskyttet inn i for eksempel nyrene.
Deler av metabolismen hos bakterier atskiller seg fra det man finner hos celler hos dyr og mennesker. Derfor er det teoretisk mulig å finne stoffer som slår ut deler av metabolismen hos bakterier, men ikke påvirker oss. Imidlertid har alle stoffer mulighet til å framkalle hypersensitivitet og allergi, og flere av dem kan i tillegg gi toksiske effekter på lever eller nyrer.
Sulfapreparater var de første vekstanalogene som ble brukt. Sulfanilamid er en vekstanalog til p-aminobenzosyre og blokkerer syntesen av folinsyre (folat). Mennesker får vitaminet folinsyre via maten, men bakterier lager sin egen folinsyre, derfor blir virkningen større hos bakterier. Halogenet fluorid (F-) er et relativt lite atom som ikke endrer de kjemiske egenskapene til stoffer nevneverdig. Derfor kan det lages vekstanaloger som inneholder fluorid f.eks. 5-fluorouracil som analog til uracil, og p-fluorofenylalanin som analog til den aromatiske aminosyren fenylanalin. 5-bromouracil er en analog til nukleinsyren thymin.
Hemmer syntese av bakteriecellevegg
Bakterieveggen er laget av peptidoglykan. Den viktigste gruppen antibiotika hører med til β-laktam antibiotika. Til denne gruppen hører penicilliner, cephalosporiner, carbapenemer, monobaktamer og cephamyciner, og felles er en β-lactamring. Penicillin lages av soppen Penicillium chrysogenum. Daptomycin påvirker celleveggen via glykopeptider og sykliske lipopeptider. Det er 3 trinn i syntesen av peptidoglykan:
I første trinn i cytoplasma blir N-acetyl-muramylpentapeptid og N-acetylglukosamin laget som UDP-derivater. D-sykloserin er et antibiotika som hemmer denne reaksjonen ved å være en analog til aminosyren D-alanin som hindrer inkorporering av D-alaninin inn i N-acetylmuramylpeptid.
Neste trinn skjer på innsiden av plasmamembranen hvor N-acetylmuramylpeptid overføres fra UDP til en lipidbærer. Bacitracin hemmer lipid fosfatase. I periplasma hos Gram-negative bakterier forlenges kjeden av peptidoglykan. Vancomycin og ristocetin virker ved å binde seg til enden av D-alanyl-D-alanin peptider, og gir hemming av enzymet transglykosylase. β-lactam antibiotika er strukturmessig lik D-alanyl-D-alanin enden i peptidoglykan og binder seg til det aktive sete på enzymet. Vancomycin virker mot slektene Bacillus, Clostridium og Staphylococcus. Det er flere enzymer som binder enheter med peptidoglykan via D-alanyl-D-alanin. Slike enzymer som binder β-lactam antibiotika kalles penicillinbindende proteiner. Eksempler på penicilliner er methicillin, ampicillin, carbenicillin etc. Isoniazid virker bare hos Mycobacterium tuberculosis ved å hemme syntesen av celleveggmateriale med mycolsyre,
Hemmer proteinsyntese
Tetrasyklin, kloramfenikol og aminoglykosider hemmer proteinsyntesen hos prokaryoter. Streptomycin isolert fra Streptomyces hemmer starten av proteinsyntesen, mens sykloheximid, kloramfenikol, puromycin og tetracyclin hemmer proteinforlengelse. Puromycin binder seg til A-sete på ribosomene. Kloramfenikol hemmer ved å blokkere dannelsen av peptidbinding. Cycloheximid påvirker bare ribosomene i Eukarya, mens kloramfenikol og streptomycin påvirker ribosomene i bakterier.
Ved transkripsjonen oversettes informasjonen fra DNA til mRNA katalysert av DNA avhengig RNA polymerase. Aktinomycin D binder seg til guanin i DNA og blokkerer transkripsjon. Rifampin (rifampicin, rifamycin) hemmer DNA-avhengig RNA polymerase ved å binde seg til β-subenheten i proteinet.
Aminoglykosider hemmer proteinsyntesen ved å binde seg til 30S subenheten i ribosomer.
Macrolider inneholder en stor laktonring festet til ett eller flere sukker, og hemmer proteinsyntese ved å binde seg til 30S subenheten på ribosomene.
På samme måte som aminoglykosider hemmer tetracykliner proteinsyntesen ved å binde seg til 30S subenheten på ribosomene.
Antibiotikaresistens
Egenskapene som gir antibiotikaresistens er ofte lokalisert til plasmider i bakteriene, og ved horisontal genoverføring forflyttes resistensen raskt mellom bakteriegrupper. De vanligste mekanismene som fremkalles antibiotikaresistens er at bakteriene lager enzymer som er istand til å inaktivere, modifisere eller kløyve molekylene med antibiotika. Det gjelder enzymet beta-laktamase, metallo-beta-laktamase og karbapenemase som bryter beta-laktamringen i penicilliner. Bakterien kan utvikle en metabolismevei som går utenom og ikke påvirkes av antibiotika, for eksempel en annen syntesevei for å lage folinsyre (vitaminet folat) som derved motstår sulfonamid-preparater eller metotrexat. Antibiotika har bindingsseter i bakteriene, men bakteriene kan endre antibiotika-reseptorene slik at bindingen hindres eller elimineres for eksempel binding av erytromycin og linkomycin. Bakteriene kan også endre permeabilitetsegenskapene i celleveggen slik at antibiotika ikke slipper inn i bakteriecellene. Det finnes også antibiotika som påvirker syntesen av DNA og RNA i bakteriene.
Etter oppdagelsen av antibiotika ble det brukt hemningsløst , ukritisk og i enorme mengder, ikke bare til å behandle mennesker og husdyr, men også innen havbruk og fiskeoppdrett, og som et middel som gir økt vekst hos kyllinger, gris og andre husdyr. Noen typer antibiotika blir forbeholdt husdyrbruk, men den kjemiske forskjellene til dem som brukes på mennesker er ikke altfor stor. Bruk av feil type antibiotika, virkningsløs bruk av antibiotika mot virussykdom, kravfulle foreldre som forlanger antibiotikabehandling av sitt syke barn, vanning av grønnsaker med kloakkvann er aspekter ved det hele. Kjenner man litt til evolusjonslæren og Darwin (Darwin-Wallace evolusjonsteori) vet man at dette må ende galt. Når antibiotikafølsomme populasjoner med bakterier utsettes for antibiotika vil de fleste dø, men ikke alle, og de som overlever har en velegnet feno- og genotype som sikrer overlevelse. Resistensgenene blir mer og mer dominerende i alle bakteriepopulasjoner, som blir utsatt for antibiotikaseleksjonspress.
På slutten av 1960-tallet og utover kommer det skremmende rapporter om bakteriestammer som tåler antibiotika. Penicillin-resistente gonorrébakterier (Neisseria gonorrhoeae) (PRNG), methicillin-resistente gule stafylokokker (Staphylococcus aureus) (MRSA), penicillin-resistent Haemophilus influenzae (PRHI), vancomycinresistente enterokokker (VRE) og multiresistente tuberkulosebakterier (Mycobacterium tuberculosis) (MDR-TB). Det dukker opp antibiotikaresistente stammer av tarmbakterier (enteriske). Resistente kolerabakterier (Vibrio cholerae), resistente tarmbakterier Escherichia coli, Clostridium, multiresistent Salmonella typhimurium, Pseudomonas aeruginosa, Shigella med shigellatoksin. Flere av bakteriene lager i tillegg mer giftige toksiner (endotoksiner) enn de opprinnelig inneholdt. Samtidig med at stadig flere resistente bakteriestammer viser seg klarer ikke den farmasøytiske industriens produksjon av nye typer antibiotika holde tritt med resistensutviklingen. Sykehusinfeksjonene (nosokomial smitte) florerer blant pasienter som allerede i utgangspunktet er svekket, forårsaket av Klebsiella pneumoniae som gir lungebetennelse og blodforgiftning, cytotoksiske gule stafylokokker, og Streptococcus pneumoniae, eller Enterococcus faecium. Høy befolkningstetthet, dårlige sanitærforhold, dårlig drikkevann, flyreiser, folkevandringer, fattigdom, ikke-hygienisk behandling og lagring av matvarer, og medisinsk kosmetisk- og tann- ”turisme” bringer de resistente bakteriene raskt over hele Jorden. I mange land er det ikke reseptplikt for antibiotika, slik at hvem som helst som ikke føler seg frisk kan kjøpe og bruke det. Plasmider spres raskt mellom bakteriestammer.
Det er ikke bare bakterier som kan utvikle resistens mot legemidler. Det gjelder bl.a. HIV-virus, men behandling med trekomponentcoctailen som påvirker flere metabolismetrinn samtidig gjør at det tar noe lenge tid før resistent virus utvikles
Mer om bakterier og virus
Litteratur
Wikipedia