Det finnes fossile eukaryoter som er ca. 2.1 milliarder år gamle. Prokaryotene er delt i to fundamentalt forskjellige organismegrupper, arkebakteriene (Archaebacteria) og de ekte bakteriene (Eubacteria). Eukaryotene som omfatter alle dagens organismer med ekte kjerne, er i fylogentisk slektskap med arkebakteriene, med utviklingsmessig splitting fra disse for ca. 3.5 milliarder år siden. Fotosyntese utviklet seg for 3.3 milliarder år siden, med anaerobe fotosyntetiske bakterier. Man antar at endosymbiose mellom grønne ikke-svovelbakterier (med fotosystem I og Calvinsyklus) og grønne svovelbakterier (med fotosystem II og CO2-fiksering med revers Krebssyklus) ga opphav til eukaryote fotosyntetiserende celler som brukte vann som elektron- og protonkilde og utskilte oksygen, slik man finner det hos blågrønnbakteriene. For 2.8-2.4 milliarder år siden finnes aerob metabolisme med de første mitokondrier.
Plantene har utviklet seg fra grønnalger og det var flere fordeler og utfordringer på land. Det er lettere å få tak i karbondioksid på land pga. høyere konsentrasjoner, og større diffusjonshastigheter av gasser i luftfase enn i væskefase. Det blir mer lys tilgjenglig. Vinden kan spre sporer og frø. Det blir værre å få tak i næringsstoffer, spesielt hvis man mangler røtter. Landplantene mister lett vann, og måtte kunne tilpasse seg tørke. Fra tiden før det ble dannet planter med frø måtte plantene ha vann tilgjengelig for at sperm skulle kunne svømme over til egget. Frøene frigjorde plantene fra vannet i befruktningen og de kunne kolonisere tørt land. Oksygen i atmosfæren la grunnlag for nitrat som er et viktig næringsstoff for plantene. Lufta har mindre tetthet enn vann og landplantene trengte en stilk eller stamme forankret til jorda med et rotsystem. De første plantene manglet røtter og blad.
Grypania er den eldste kjente spiralformet alge og er funnet i lagdelte jernformasjon i N-Michigan i USA. I Huntington formasjonsjonen i arktisk N-Canada finnes fossiler av rødalger som ligner på dagens Bangia. Flercellete grønnalger funnet på Svanbergfjellet på Spitsbergen ligner på dagens Cladophorales med filamentformet thallus og sylindriske celler festet til hverandre med septalplater.
Under evolusjonen fikk landorganismene en reproduksjon som var mindre avhengig av vann i reproduksjonen. Det er funnet sporer fra planter fra Ordovicium for ca. 450 millioner år siden og hele planter fra tidlig i Devon for ca. 408 millioner år siden. På denne tiden har man observert meiose av en diploid celle som ga en tetrade.
Landplantene har sannsynligvis utviklet seg fra grønnalger (Chlorophyta) f.eks. fra kransalger (Chara) tilhørende klassen Charophyceae. Både landplanter og grønnalger inneholder fotosyntesepigmentene klorofyll a og b i kloroplaster, og cellulose og hemicellulose i celleveggene Begge har stivelse som opplagsnæring. For planter som har bevegelige celler er flagellapparatet det samme som hos grønnalger. Det finnes også biokjemiske likhetspunkter. Glykolat oksidase finnes hos planter og kransalger (Chara og Nitella), mens grønnalger for øvrig bruker glykolat dehydrogenase, og på denne måten resirkuleres karbon som ellers ville ha blitt tapt i fotorespirasjon. kobber-zink (CuZn) superoksid dismutase (SOD) finnes både hos Chara og planter. Arter i Charophyceae som Chara, Spirogyra og Coleochaete har sporopollenin i zygoten. Sporopollenin finnes i pollen fra planter.
Blader utvikles fra mikrofyll til makrofyll
Bladene utfører fotosyntese, og er torneformete utvekster som under evolusjonen blir til nåleformete blad med en bladnerve med ledningsvev. Disse kan etter hvert utvikle seg til større bladplate med mer nervatur. Blader kan også ha utviklet seg av flate dikotome greiner som har vokst sammen.
Mikrofyll er små utvekster (blad) rett ut fra stengelen med en ugreinet streng med ledningsvev. Mikrofyll finnes hos kråkefotplantene. Bower presenterte i 1935 en Enation-teori (l. enatus - vokser ut fra) hvor en tornelignende utvekst kunne bli koblet til en enkel sentral streng med ledningsvev. Slike tornete utvekster kan man finne hos Sawdonia og andre arter i Zosterophyllum. Torneutvekstene fikk først ledningsvev ved basis, som hos Asteroxylon. Seinere vokste ledningsvevet ut i hele mikrofyllet som hos kråkefot (Lycopodium). Telomteorien til Zimmermann fra 1952 sier at sammenvoksing av dikotome gaffeldelte greiner etter hvert ga en flat bladplate kalt megafyll (eufyll) som inneholdt ledningsvev i form av en sifonostele eller eustele. Mellom de gaffeldelte greinene som lå i en plan flate ble det dannet vev. En av greinene blir dominerende og det dannes en plan bladflate optimalisert for å fange opp sollys. Megafyll er det mer komplekse blad som danner bregnefronder, og finnes hos sneller og gymnospermer. Megafyll utvikles i Devon hvor konsentrasjonen av karbondioksid i lufta blir lavere.
Tørketålende hud med gassutveksling
Den økologiske nisjen til alger i tidevannssonen krevde at algene måtte tåle uttørking ved fjære sjø. Det er sannsynlig at de første plantene lignet alger f.eks. Coleochaete som vokser på jord og Parka med thallus med pseudoparenkym, som vokste utover underlaget med sporangier fra overflaten. Det er rimelig å anta at ikke-vaskulære planter som moser, levermoser og nålkapselmoser var de første til å bli terrestre og kolonisere land. Overgangen fra vannliv til landliv krevde store endringer i morfologi og anatomi. Det ble behov for en vanntett epidermis og kutikula med gassutveksling gjennom porer (spalteåpninger). Plantene trenger karbondioksid fra lufta, men taper samtidig vann. Dyrene utviklet lunger.
Transportsystem og styrkevev
Planter og dyr som lever neddykket i vannmettede omgivelser kan ta opp vann fra omgivelsene. Plantene lager strukturer som tar opp vann fra jorda. Det fantes rotlignende systemer hos alger som kunne ta over denne funksjonen. Dyr som kan forflytte seg må ha en vannkilde i nærheten som de kan drikke av. Dyr har et sirkulasjonssystem som vannet kunne forflyttes i kroppen med.
Vann er et mye tettere medium enn luft. Under vann trenger ikke en plante støtte i reisverket for å kunne stå oppreist sammenlignet med luft. For å klare dette har de terrestre dyrene et skjelett. En oppreist plante krevde styrkevev med lignin og et system for langdistansetransport, vaskulært vev bestående av xylem (vedvev) som frakter vann og næringssalter fra jorda opp i planten og floem (silvev) som frakter fotosynteseprodukter til den delen av planten der det er størst behov.
Høyere planter krevde et mer effektivt transportsystem med utvikling av en stele. Protostele hos de første plantene som kråkefotplanter (lycopsider) og snelleplanter (sphenopsider), bestående av en solid streng med xylem omgitt av floem, og ytterst en endodermis. En mer avansert stele var sifonostele som består av en sylinder med marg som er omgitt av xylem og floem, og utenfor floemet en endodermis. Denne finnes hos bregner (filicopsider), kråkefotplanter (lycopsider) og sneller (sphenopsider). Eustele består av atskilte strenger som inneholder xylem og floem, atskilt av parenkym. Ledningsstrengene kan ligge i en sirkel eller spredt over hele stengeltverrsnittet hos urtaktige planter.
Plantene spiller en viktig rolle for vannsyklus på jorda. En sentral stele med vedvev (xylem) omgitt av silvev (floem) var nok for lave vaskulære planter, men etter hvert som plantene ble høyere krevdes mer avansert ledningsvev. Først ble det sentrale vaskulære vevet fylt med en sylinder med parenkym. Det primære vaskulære vevet er ikke treaktig. Ved (sekundært xylem) er treaktig og gir mer styrke. Tre mest vanlig å finne i fossilt materiale siden det er mer resistent mot nedbrytning og hardere.
Reproduksjon på land
Algene hadde sporer som kunne spres med vannstrømmer. De første landplantene hadde sporer som kunne spres med vann og vind. Sporene måtte også kunne tåle uttørking. Reproduksjon hos vanndyr og alger går ut på å utveksle gameter under vann og det skjer en ytre befruktning. De viktigste forandringene som skjedde med landplantene var utvikling og spesialisering av de reproduktive strukturene, tilpasset et stadig tørrere habitat etter som de koloniserte mer og mer av tørt land.
Generasjonsveksling mellom sporofytt og gametofytt
Algene har en diploid sporofytt og en haploid gametofytt som frigir hannlige og hunnlige gameter. Ved befruktningen smelter gametene sammen (fusjon) og gir en zygote som vokser opp til en ny diploid sporofytt, en generasjonsveksling. Gametofyttgenerasjonen kan leve på sporofyttgenerasjonen. Sporofyttgenerasjonen kan også lage en uavhengig gametofytt. Hos karplantene (vaskulære planter) er sporofytten størst og mest synlig dominerende. Plantene veksler mellom en flercellet diploid fase (sporofyttfase) og en haploid fase (gamtofyttfase). Hos mosene (bryofyttene) er det den haploide gametofyttfasen som er dominerende i størrelse.
Ved seksuell reproduksjon, som er vanlig hos eukaryotene, finnes det to kjerner kalt gameter som kommer fra hver av foreldrene og som smelter sammen og danner en ny kjerne (zygote). Gametkjernene finnes i en enkelt celle, kalt henholdsvis sperm og egg. Ved definisjon er hannen den som bevegelige gameter. Når gametene smelter sammen blandes to slags kromosomer, en fra hver av foreldrene og kjernen kalles diploid. Gametkjernen med bare ett sett kromosomer kalles haploid. Haploide celler dannes under reduksjonsdelingen (meiosen).
Bregnene slipper ut haploide sporer laget i en meiose i den diploide planten kalt sporofytt. Sporene kan utvikle seg til små planter som ser helt forskjellig ut fra morplanten og kalles gametofytter fordi de lager gameter som kan være bevegelige (sperm) eller ubevegelige (egg). Hva er fordelen med skifte mellom sporofytt og gametofytt ? Plantene trenger ikke nær kontakt med en artsfrende, idet en spore kan havne på et sted med gode vokseforhold. Plantene er stasjonære og kan ikke oppsøke en seksualpartner. De første gametofyttene levde i et fuktig miljø fordi sperm måtte svømme over til egget. Vi kan si at gametofytten egentlig er en vannplante. Sporofytten kan derimot leve under tørrere betingelser. Akkurat som amfibiene var plantene i starten avhengig av vann i reproduksjonen. Hos mosene er det gametofytten som dominerer og sporofytten er uanseelig. Hos bregner blir gametofytten mikroskopisk og atskilt fra sporotfytten. Hos frøplantene blir hunngametofytten værende inne i sporofytten og det hannlige gametofytten som er pollenkornet spres med vind, vann eller insekter.

Evolusjonsmessig opprinnelige og avledete egenskaper hos planter
Homospore planter
Planter som lager to morfologiske forskjellige sporer kalles heterospore, og de som lager bare en type sporer kalles homospore.
Sporene lages i sporangier som resultat av meiose i sporofyttgenerasjonen. Differensieringen av dem skjer i spesielle flercellete strukturer kalt sporangier (ent. sporangium). En homospor plante har homosporer som når de spirer gir en haploid flercellet gametofytt. Frittlevende gametofytter kan leve i symbiose med sopp. Soppene, i form av mykorrhiza, utviklet seg sammen med landplantene. Sporene vokser opp til en klorofyllfri underjordisk eller overjordisk grønn gametofytt med fotosyntese. Det skjer altså ikke noe befruktning eller kjønnslig utveksling før gametofytten vokser opp. Gametofytten er alltid haploid og kjønnscellene kan derfor lages ved vanlig celledeling (mitose). Gametene lages i spesielle organer på overflaten av gametofytten, kalt gametangier, hvor egg og spermceller omgis av et hylster. Kammere som gir eggceller kalles arkegonier, og de som gir spermceller kalles anteridier. Det lages et egg i hvert arkegonium. Befruktningen skjer ved at spermatozoider svømmer ut av anteridene gjennom en åpning (operculum), og i en vannfilm forflytter de seg over til halsen til arkegoniet gjennom en kanal til egget. Den første cellen som dannes etter befruktningen er en diploid zygote som ved celledelinger vokser til et embryo som utvikler seg i arkegoniekammeret omgitt av gametofyttvev som sørger for næringstilførsel til den voksende embryo. Etter hvert vokser sporofytten ut av gametofytten, danner røtter og fotosyntetiserende blad og danner en selvstendig sporofyttgenerasjon.
Hos en del moser og bregner er anteridier og arkegonier på samme gametofytt. Hvis gametofytten som bærer anteridier og den andre som bærer arkegonier er forskjellig kalles den som danner anteridier mikrogametofytten, og den som danner arkegonier kalles makrogametofytten. Alle mosene er homospore. Mosene finnes på fuktige steder, og i Arktis og Antarktis er de de meste vanlige plantene. Gametofytten hos mosene er grønn og fotosyntetiserende i forhold til sporofytten som får næring direkte fra gametofytten. Tuer med moseplanter gir større mulighet for at svømmende spermceller på en fuktig overflate skal kunne svømme over til egget. Mosene vokser på fuktig skogbunn.
Det skjedde en overgang fra homospore til heterospore planter. I Silur var det homospore planter. I mer spesialiserte vaskulære planter er gametangiene forsvunnet.
Heterospore planter
I nesten alle planter kommer gametofytten fra to forskjellige typer sporer (heterosporer) kalt mikrosporer og megasporer. Megasporer (100-200 mikrometer (μm) i diameter) og mikrosporer (diameter mindre enn 50 μm), ble første gang funnet i Devon. Hvis en plante lager haploide mikro- og megasporer kalles sporangiene henholdsvis mikrosporangier og megasporangier. Til forskjell fra de homospore plantene lager de heterospore to typer gametofytter, henholdsvis mikrogametofytt og megagametofytt. Celledelinger i mikrosporen gir en mikrogametofytt med anterider som lager sperm. Megasporer danner en megagametofytt med arkegonier som inneholder egg. En annen forskjell mellom homospore og heterospore er at gametofytten lages på utsiden av sporeveggen ved spiringen hos de homospore, men på innsiden av sporeveggen hos de heterospore. Når egget er modent åpnes en spalte slik at spermceller kan komme fram til egget.
Heterospori er forløperen for frø i gymnospermer og blomsterplanter.
Mosene - de første landplanter
Bryofytter er (Bryophyta) er ikkevaskulære planter og omfatter moser, levermoser og nålkapselmoser.
Ekte moser (Bryophyta)
Mosene har en vekstform hvor hele planten danner lave tuer og de foretrekker fuktige habitater. Det er ikke det samme behov for langdistansetransport hos moser, og mosene mangler avansert ledningsvev med xylem (vedvev) med lignin, samt floem (silvev). Transporten skjer ved diffusjon og osmose, og det begrenser størrelsen på mosene. Hos de mest avansert mosene som bjørnemose er det ledningsvevsceller kalt hydroider og leptoider (gr. leptos - tynn, fin). Det finnes ca. 9000 nålevende arter moser. Moseplanten er ikke-vaskulær og består av en stengellignende struktur med bladlignende utvekster bestående av ett lag udifferensierte celler, unntatt ved midtnerven. Moser er festet til bakken med rhizoider. Strukturene i moseplanten er ikke homologe med rot, stengel og blad hos vaskulære planter.
Mosene har et flercellet gametangium, og sterile celler omgir og beskytter gametene. Algene har encellet gametangium. Det befruktede egget gir en zygote som vokser til et beskyttet flercellet embryo. Gametofyttgenerasjonen er den haploide fasen som gir haploide gameter. To gameter fusjonerer og danner sporofyttgenerasjonen som lager haploide sporer via meiose. Den haploide gametofytten lager hannlige gametangier (anteridier) som lager sperm, samt hunnlige gametangier (arkegonier) som lager egg. Gametangiene befinner seg på toppen av moseplanten. Mange moser har atskilte hann- og hunnmoser, men hann og hunn kan også befinne seg på samme mose. Spermceller med flagell er avhengig av vann eller insekter for å komme over til arkegoniene hvor sperm svømmer inn arkegoniehalsen som skiller ut spermtiltrekkene stoffer. Sporofytten vokser opp på toppen av den hunnlige gametofytten. Den første tiden er sporofytten grønn og utfører fotosyntese, deretter størker den ut. En fot fester gametofytten og består i tillegg av en seta (stilk) og en kapsel med sporogene celler. Kapslen kan være dekket av en kopp (kalyptra). Sporofytten har sporogene celler (sporemorceller) som ved meiose danner sporer. En spore vokser opp til et filament (protonema) som ligner en alge. Protonema danner knopper og vokser opp til en grønn gametofytt. Mosene er med å hindrer erosjon.
Levermoser (Hepatophyta)
Det er ca. 6000 nålevende arter levermoser som lever på fuktig jord. Levermosene har et flatt thallus med rhizoider. Noen har bladlingende bald med ett lag udifferensierte celler, og en "stengel" med rhizoider. Levermosene mangler spalteåpninger (stomata), men har porer på overflaten. Den seksuelle formeringen skjer via arkegonier og anteridier på en haploid gametofytt. Hos noen levermoser finnes gametangiene på arkegoniforer og anteridioforer. En regndråpe fører sperm over til arkegoniet. Zygoten danner et embryo som vokser opp til en sporofytt som er festet på gametofytten. Sporofytten består av fot, seta og kapsel med sporogene celler som via meiose gir sporer. Vev fra arkegoniet omgir kapselen. En spore spirer til en ungt gametofytt og det finnes hannlige og hunnlige gametofytter. Aseksuelt kan levermosene formere seg fra groknopper (gemmae, ent. gemma) i koppformete strukturer på thallus. Utvekster fra thallus kan strekke seg og danne et uavhengig thallus. Det er funnet rester av levermoser i 400 millioner år gamle kulleier.
Nålkapselmoser (Anthocerophyta)
Det finnes ca. 100 nålevende arter nålkapselmoser som lever i forstyrrete habitater. Thallus ligner det man finner hos levermosene, men de har bare en stor kloroplast i hver celle. Moser og levermoser har mange kloroplaster per celle. Etter befruktning vokser det opp en nålformet sporofytt. En gametofytt lager flere sporofytter. Det skjer meiose i hver sporangium. Sporene kommer ut i spalte på toppen av sporofytten som fortsetter å vokse fra basis.
Karsporeplanter - frøløse vaskulære planter (Pteridophyta)
Karsporeplantene utviklet seg for 420 millioner år siden fra midten av den geologiske tidsperioden Silur.
Karsporeplantene har to typer blad: mikrofyll og makrofyll. Hos bregnene er det en flerårig sporofytt som er dominerende i størrelse, i motsetning til mosene. Bregnene har rhizom, og blader som kalles fronder. Tidligere lette man etter "bregnefrø", men man fant ikke noe. Derimot observerte man støvet fra svarte eller brune streker (nå kalt sori) under bregneblad. Linné mente at "pulveret" så ut som pollen. Malpighi og Grew undersøkte "pulveret" i mikroskop. Sporer dannes på områder på frondene som utvikler sporangier. En samling sporangier kalles sori (ent. sorus), og hvert sori inneholder hundrevis av kuleformete sporangier med en tynn stilk. Det er ca. 64 sporer i hver sporekapsel, og det går et tykt segmentert bånd, annulus, omkring kapselen som skyter ut sporene som en katapult. Hos noen arter er hvert sori dekket av et indusium som beskytter sporangiene under. Sisselrot mangler indusium, og Adiantum har et falskt indusium dannet fra kanten av fronden. I sporangiet er det sporogene celler som via meiose danner haploide sporer. Lindsay oppdaget i 1794 at det var mulig å få nye bregner fra Abregnefrø@, og han sendte slike Abregnefrø@ til Joseph Banks i London. Men hvis støvet under bladene var Afrø@, hvor var så pollen ? En spore spirer og danner ved mitose en gametofytt (prothallus). I 1844 oppdaget Karl von Näegeli at på undersiden av de grønnfargete tynne membranformete flate hjerteformete skjellene (bregneprothalliene) som kom fra sporene så var det runde vorter eller papiller. Samme type papiller var blitt funnet hos moser og levermoser. Nägeli fant at i noen av papillene var det mørke spiralfilamenter, og når våte papiller brast kom det ut spiralfilamenter som beveget seg mot en annen type papiller. Hos karsporeplantene vokser det opp en gametofytt fra en spore. En spore er en celle. Gametofyttfasen er en seksuell generasjon som lager kjønnsceller. Sporofyttfasen kommer fra en zygote.
Bregnegametofytten (prothallus) mangler ledningsvev og er festet til bakken med rhizoider, og produserer både arkegonier og anteridier på undersiden. Arkegoniet har en eggcelle. Spermceller med flagell svømmer fra et nærliggende prothallus over til arkegoniet, ned arkegoniehalsen og inn til egget. Egg og sperm fusjonerer og det dannes en zygote. Zygoten danner en sporofytt. Sporofytten vokser opp på gametofytten, prothallus dør deretter og det er dannet en selvstendig sporofytt.
Bregnene står evolusjonsmessig nærmere frøplantene sammenlignet med kråkefotplantene. Lykofyttene er monofyletiske (Lycophytina) og bregner og frøplanter inngår i gruppen med megefayll (Euphyllophytina) (Schneider et al. 2004).
Psilofytter (Psilophyta)
12 nålevende arter psilofytter som mangler røtter og blad, og sporehuset er inndelt i rom. Psilotum som vokser i tropiske og subtropiske områder har dikotom forgreining av en stengel med fotosyntese og sporangier samlet i synangier. Sporene er bønneformete og ikke runde som hos de første landplantene. Det dannes små runde sporangier i enden av stengelgreinene. Haploide sporer spirer til et haploid ikke fotosyntetiserende prothallus som vokser under jorda sammen med mykorrhiza. Psilotum står nær Tmesipteris som vokser i Australia som en epifytt på trebregner. Psilofytter og sneller er egentlig bregner.
Sneller (Equisetopsida) (l. equus - hest; seta - bust, kort stivt hårstrå)
15 nålevende arter sneller som vokser I fuktig habitat. Minner om halen på en hest (eng. horsetails). Har rot, hul stengel og kranser med små blad (redusert megafyll) i ringer festet til nodier. I enden av stengelen dannes en konglelignende strobilus. (-opsida - klasse). Snellene er egentlig bregner, og i slekt med arter i kongsbregnefamilien (Osmundaceae, orden Osmundales) og Marattiaceae (orden Marattiales).
Kråkefotplanter/lykofytter (Lycophyta ) (gr. lycos - ulv; pous - fot)
Lykofyttene har sporangier på oversiden av blad (mikrofyll) eller i aksilen mellom blad og stengel. Dette er forskjellig fra bregnene som har sporer på undersiden av blad. Dessuten har lykofyttene enkle blad (mikrofyll) med en nerve og uten bladgap, mens bregnene har fronder (makrofyll). Bladgap vil si et gap i det vaskulære vevet der hvor bladet går ut fra stengelen, og gapet er fylt med parenkymceller. Nålevende lykofytter er små planter med skjellignende blad (mikrofyll). Dagens kråkefotplanter er små planter som dvergjamne (Selaginella) i ordenen Selaginnales, kråkefot (Lycopodium) i ordenen Lycopodiales og brasmegras (Isoetes) i ordenen Isoetales. Brasmegras er en vannplante med kormuslignende jordstengel, hvorfra det kommer opprette smale torne- eller grasformete blad i spiral. Modifiserte blad kalt sporofyll bærer enten makrosporangier eller mikrosporangier på oversiden (adaksialt) av bladbasis. Bladene hos brasmegras har fire luftkanaler (aerenkym) som frakter karbondioksid fra bunnsedimentene opp i bladet, og på den måten unngår de problemet med den lave diffusjonshastigheten for gasser i vann sammenlignet med luft. Både Selaginnales og Isoetales inneholder heterospore arter. Tidligere var kråkefotplantene trelignende planter, bl.a. skjelltrær (lepidodrendrider). Sporene hos lykofyttene spirer i mørke og blir til underjordiske prothaller uten klorofyll, og de lever i symbiose med sopp.
Selaginella er heterospor med en strobilus med to typer sporangier:
1. Makrosporangier som inneholder megasporocyter (megasporemorceller) som ved meiose danner store megasporer. Megsporene danner en hunnlig gametofytt med egg i arkegonier, men dette skjer innenfor sporeveggen, dvs. det dannes ingen frittlevende gametofytt.
2. Mikrosporangier som inneholder mikrosporocyter (mikrosporemorceller) som ved meiose danner mikrosporer. Mikrosporene danner en hannlig gametofytt med spermceller i anteridier. Hvert anteridium lager mangle spermceller.
Etter et spermcellene har befruktet egg vokser den unge sporofytten ut av den hunnelige gametofytten.
Frøplantene (Spermatophyta)
Det skjedde en overgang fra heterosporer til frø hos frøplantene. Hos bartrærne var det ikke bare forskjell i størrelsen på sporene, men de ble atskilt på planten, med mikrosporer og makrosporer i henholdsvis pollenkongler og frøkongler. En fåcellet mikrogametofytt (pollenkorn) inneholder spermceller i en pollenslange som brukes til å befrukte frøemnet. Det skjer abort av tre av megasporene og en megaspore som overlever, og gir utvikling av et frøemne. Sporangiet blir i tillegg omgitt av integumenter som gir utvikling av frøskall. I spissen av integumentene er det en åpning kalt mikropyle som fører inn til et pollenkammer. Hos gymnospermene skiller frøemnet ut en pollinderingsdråpe fra nedbrytning av integumentene, som pollen spredt med vinden kan feste seg i. Pollen blir overført direkte til frøemnet hos gymnospermene, men via arr og griffel hos angiospermene. Hos heterospore og homospore kommer sporene ut av sporangiene, men hos frøplantene blir den funksjonelle megasporen holdt innenfor sporangieveggen
Pollen er forskjellig fra sporer ved hvordan de avgir gameter. Mens mikrosporene gir gameter med flagell som svømmer til arkegoniet fra proksimal ende, vokser pollenslangen ut fra distal ende i pollenkornet.
Hos planter skjer meiosen i spesielle celler kalt sporemorceller og resultatet er haploide sporer, de første cellene i gametofyttgenerasjonen. Sporene fusjonerer ikke som gameter, men deler seg ved mitose og gir en flercellet haploid gametofytt. Gametene (egg og spermceller) lages av gametofytten ved mitose. Når gametene fusjonerer danner de en zygote, den første cellen til sporofyttgenerasjonen.
Hos de dekkfrøete blomsterplantene (Angiospermophyta) består gametofytten av noen få celler (embryosekken og pollenkornet).
Frøemnet med embryosekk tilsvarer den frittlevende gametofytt hos bregner (prothallus). Embryosekken har åtte kjerner dannet ved tre mitotiske kjernedelinger av en megaspore. Megasporen kommer fra en meiose fra opprinnelige fire megasporemorceller. Haploid gametofyttgenerasjon etterfulgt av en diploid sporofyttgenerasjon. Hos plantene forekommer egne hann- og hunnplanter selv om tøkjønnete er vel så vanlig. Sakkat pollen (l. saccus - sekk) har luftsekker som hjelper til under vindspredning. Pollen kunne ha med lang spalte (sutur), og klebrig pollen kunne feste seg. Pollen har en ytre vegg kalt eksine, med tektum og kolumella, og en indre intine. Angiospermene har tradisjonelt vært delt i en- og tofrøbladete planter. Nå delt i to monofyletiske grupper monokoter, og eudikoter som omfatter de fleste, men ikke alle tofrøbladete, og har pollen med tre spalter (aperturer). Pollen med tre spirefurer har lettere for å spire på arret, siden dette øker sannsynligheten for at en av furene vender ned mot arret. Den andre gruppen tofrøbladete er magnoliidene med pollen med en apertur. De tofrøbladete plantene utviklet seg først. Enfrøbladete planter utviklet seg fra tofrøbladete planter for 135-75 millioner år siden.
Dobbel befruktning hvor både sentralkjerne, som danner triploid endosperm, og eggcellen blir befruktet med hver sin spermkjerne, sikrer at bare de frøemnene hvor egget er blitt befruktet fylles med opplagsnæring. Etter befruktning blir frøemne (megasporangium) og omgivende vev til et frø. Integumentene (sporofyttvev) som omgir megasporangiet blir til frøskall. Dobbel befruktning ga frø med opplagsnæring som lettere kunne etablere seg. Frøene er attraktive for dyrene som deltar i spredning av frøene samtidig som opplagsnæringen og den tilhørende frukten ga grunnlag for videre utvikling av pattedyrene.
Gymnospermene (nakenfrøete) har ingen fruktknutevegg og frøet blir liggende på et kongleskjell.
Angiospermene (dekkfrøete) har en fruktknutevegg som omgir fruktknuten som inneholder ett eller flere frøemner. Angiospermene lager frø inne i en frukt, som gir beskyttelse under frøutviklingen og frukten kan delta i spredning av frøene. Det er generasjonsvekseling, sporofytten dominerer i størrelse, gametofytten er blitt mikroskopisk og er fullstendig avhengig av sporofytten, og befruktningen er mindre avhengig av vann. Drimys piperata har et bladformet fruktblad. Både gymnospermer og angiospermer er heterospore.
Plantene har utviklet seg over mer enn 450 millioner år fra flercellete grønnalger. Plantene ble terrestriske og kolonisert land i overgangen Silur til Devon. For 300 millioner år dannet trær store skoger. Gras hadde sin opprinnelse for ca. 65 millioner år siden. Noen arter er stabile og morfologisk like i millioner av år f.eks. Ginkgo, bregner og Cycas, og kan eksistere i små populasjoner. Evolusjon er små forandringer over tid, men det kan skje rask artsdannelse avbrutt av stabile perioder (punktuert likevekt). 266.000 arter planter er kjent. Det er ca. 250.000 -300.000 vaskulære planter og 16.000 moser.
Vaskulære planter (karplanter) (Trachaeophyta) omfatter karsporeplanter og frøplanter. Mosene og de vaskulære plantene fikk atkskilte utviklingslinjer for lang tid tilbake. Mosene levde på fuktige steder og hadde ikke tilpasset seg fullstendig til et liv på land. Mosene hadde ikke nok styrkevev og nok velutviklete transportsystem, og manglet røtter for å kunne vokse på tørrere habitater. Den andre store utviklingslinjen går fra planter som produserte sporer til de som laget frø. De første vaskulære plantene laget bare sporer. Frø var en usedvanlig vellykket spredningsenhet og inneholdt en flercellet liten plante (embryo) omgitt av opplagsnæring og beskyttet av et frøskall. Frø har også betydning for utvikling av menneskelig sivilisasjon. Frøene kunne samles og lagres tørt.
Bartrær (Coniferophyta)
Bruker furu (Pinus) som eksempel. Bartrærne er eviggrønne, unntatt lerk (Larix), har nåleformete blad, harpiks og trakeider, men mangler vedrør. De er monoike med atskilte hann og hunn på forskjellig sted på samme plante. Megasporer og mikrosporer dannes på forskjellige kongler. Pollenkonglene med sporofyll er en bladlignende struktur med sporganier på undersiden. Ved basis av hvert sporofyll er det to mikrosporangier med mange mikrosporocyter (mikrosporemorceller). Hver mikrosporocyt danner ved meiose fire haploide mikrosporer. Pollenkorn består av generativ celle og vegetativ celle (tubecelle som danner pollenslange), og to celler degenererer. To luftsekker gir oppdrift av pollenkornet. Frøkonglen (hunnkonglen) er et modifisert greinsystem. Hvert kongleskjell har to frøemner (magasporangier) på overflaten. I hvert megasporangium gir meiose av en megasporocyt fire haploide megasporer. Tre av megasporene desintegrerer, mens den fjerde deler seg mitotisk og danner en hunnlig gametofytt som inneholder et egg i et arkegonium. Frøemne produserer en klebrig dråpe hvor pollen lander under pollenspredningen om våren. Pollenkonglen forsvinner. Pollen utvikler en pollenslange som vokser gjennom megasporangiet til egget i arkegoniet. Pollenkorn med pollenslange er den hannlige gametofytten. Det er den vegetative cellen (tubecellen) som danner pollenslangen. Den generative cellen kommer inn i pollenslangen, deler seg og danner en stilkcelle og en kroppscelle. Kropsscellen deler seg og danner to ubevegelige spermceller som blir frigitt nær egget. En av spermcellene fusjonerer med egget og danner en zygote. Den andre spermcellen desintegrerer. Embryo bestående av embryo, embryoskudd og embryorot og flere kimblad er begravet i den haploide hunnlige gametofyttvevet som blir næringsvev i det modne frø. Integumenter danner frøskall som omgir embryo og næringsvevet. Det går lang tid fra pollinering til befruktning. Ved pollineringen er hunnkonglen umoden, og det har ikke skjedd meiose av megasporocyten. Etter at megsporen er dannet mer enn et år i forveien blir det dannet et egg i arkegoniet. Imens vokser pollenslangen sakte gjennom megasporangiet til arkegoniet. Befruktning skjer ca. 15 månder etter pollinering. Frøutviklingen tar flere måneder. Den hannlige gametofytten lager ikke anteridier og spermcellene er ikke avhengig av vann for å komme fram til egget. En ikkebevegelig sperm blir fraktet via pollenslangen fram til egget, en tilpasning til liv på land.
Konglepalmer/cycadeer (Cycadophyta)
Konglepalmer utviklet seg i Trias for 248 millioner år siden. 140 nålevende arter i tropiske og subtropiske områder, men mange er utdødde.Har sammensatte blad og en tønneformet stamme. Dioike planter med frøkongler på hunnplanter og pollenkongler på hannplanter. Sperm har en hårlignende flagell, en rest fra den tid da sperm svømte fra anteridium til arkegondium. Nå er det vind eller insekter som frakter pollen som vokser ut i en pollenslange.
Ginkofytter (Ginkgophyta)
Ginkgo biloba er den eneste gjenlevende arten gingkofytter. Har vifteformete blad med høstfarger. Har spiselige frø og dyrkes I Ø-Kina. Treet er herdig og tåler luftforurensning og plantes i nåtidens byer. Arten er dioik med egne hann og hunntrær, men det er hanntrærne som plantes, fordi hunntrærne har frø med et kjøttfylt ytre som gir en ubehagelig lukt av harsknet smør (smørsyre). Lukten kan muligens skremme vekk frøspisere. Spermceller med flageller spres med vinden. Frøene ligger eksponert. Ekstrakt fra blad brukes medisinsk.
Gnetofytter (Gnetophyta)
Består av tre slekter med 70 arter gnetofytter. Gnetum har eksponerte frø. Ephedra har pollenkongler i nodiene, og vokser i tørre områder. Noen Ephedra har grønn stengel og små blad. Inneholder stoffet efedrin som aktiverer hjerte og øker blodtrykket. Welwitschia vokser i SV-Afrika og har en lang pelerot. Har to blad som vokser hele tiden, men det ser ut som flere. Kongler langs kanten. Hos gnetofyttene finnes vedrørselementer. Ephedra og Gnetum har dobbel befruktning, men det dannes en ekstra zygote istedet for endosperm, og den andre zygoten desintegrerer.
Blomsterplanter (Magnoliophyta)
Blomsterplantene (angiospermene) deles i klassen enfrøbladete (Liliopsida) som omfatter gras, palmer, løk, liljer og orkidéer, og klassen de tofrøbladete plantene (Magnoliopsida). De endrøbladete plantene, idag ca. 60000 arter danner en monofyletisk gruppe med felles evolusjonshistorie tilbake til Kritt,125-113 millioner år siden). Den viktigste familien av enfrøbaldete er grasfamilien, som gir beitegras til plantespisere, men inneholder også de domestiserte kornslagene hvete, rug, havre, bygg, mais, ris, sorghum, teff, og har således hatt stor betydning for evolusjon av mennesket og dannelse av menneskesamfunn.
Hos blomsterplantene vokser fruktveggen til en frukt som beskytter frøet og hindrer uttørking når frøet vokser og sørger for spredning. Blomsterplantene med dobbel befruktning har stor reproduktiv suksess og dominerer jorda i stort antall. Brede blad samler mye sollys, og bladverket kastes av ved tørke og kulde og derfor kunne blomsterplantene også innta økologiske nisjer som var ugjestmilde en del av året. Opplagsnæring for flerårige planter finnes i jordstengel (rhizom) eller rot. Kaktus tåler tørke, stengelen sørger for vannlagring og fotosyntese, og nålene er omdannete blad som beskytter mot plantespisere.
Det går en utviklingslinje fra karsporeplanter til progymnospermer med megafyll og sekundært xylem. Archaeopteris er en progymnosperm mellom sporeplantene og frøplantene. Mikrosporene ble omdannet til pollen, og megasporangium ble til frøemne. Frøplantene utviklet seg uavhengig av hverandre og flere ganger fra Devon for 360 millioner år siden. Gingko finnes 200 millioner år siden. Progymnospermene hadde utviklingslinjer til bartrærne og en annen linje førte fram til frøbregner og ginkgofytter. Emplectopteris er en frøbregne som hadde frø fra bregnelignende blad. De eldste blomsterplantene er kjent fra Jura-Kritt for 125 - 145 millioner år siden.
Fra Silur kan man observere trakeider med veggfortykkelser og plantene kunne stå oppreist og var tilpasset tørke. Dette var de første landplantene fra Silur-Devon som var små planter. Alger og moser levde nær vannkanten før de vaskulære plantene. Det man ser etter i fossiler er ledningsvev (xylem), kutikula, sporer med en eller spalte/porer, og spalteåpninger på blad.
Man har antatt at Cooksonia, ca. halvmeter høy, var blant de første vaskulære plantene fra midten av Silur. Cooksonia hadde dikotom forgreining med sporangier med sporer i spissen. Det var ingen blader, men hele den grønne stengelen kunne utføre fotosyntese. Cooksonia manglet røtter, men var festet med rhizom, en forlengelse av stengelen.Monopodial foregreining som ble utviklet under midten av Devon var en mer avansert greiningsmønster.
Rhynia var en ca. 20 cm høy plante festet til jorda med et rhizom, var oppreist og dikotomt forgreinet med sporangier i spissen av greinene. Rhynia hadde en stele med trakeider sentralt plassert omgitt av floem (sentrark protostele) Rhynia ble funnet i Rhynie Chert i Aberdeenshire i Skotland, et sted som inneholder plantefossiler fra nedre Devon, bl.a. Aglaophyton, Asteroxylon og Horneophyton. Rhyniofyttene vokste sannsynligvis i myrer og på andre fuktige steder.
Zosterophyllum er et eksempel på en mer avansert plante som hadde runde eller ovale sporangier på små stilker lateralt på stengelen, og ikke i enden (terminalt) av stilkene som Cooksonia og Aglaophyton. Zosterophyllum fantes fra nedre Devon, hadde ikke røtter, men på stammen hadde den grønne tornelignende utvekster som var forløperen til blader. Fra Zosterophyllum kunne det gå en utviklingslinje til kråkefotplantene (lykofyttene), som hadde sporangiene på oversiden av mikrofyll eller i hjørnet mellom stengel og mikrofyll. Hos Zosterophyllum hadde også grønne greiner som kom opp fra basis, og som manglet sporangier og utførte fotosyntese. Baragwanathia fra Australiadelen av Gondwana hadde lang stengel med rhizom, og var en primitiv kråkefotplante.
I Trimerophytina forekom arter som Trimerophyton og Psilophyton. Psilophyton hadde rikere forgreining med vegetative og reproduktive greiner, var små planter og fantes i midten av Devon. Sporangiene var samlet i hoper på enden av greiner. Tornelignende utvekster fantes på stengelen. De vaskulære plante (trakeofyttene) med trakeider deles i Rhyniopsida med Rhynia og Eutracheophyta.
Eutracheophyta deles i:
1) Lycophytina hvor sporangiene er plassert lateralt på en bladløs stamme f.eks. Zosterophyllum og Baragwanathia (Zosterophyllopsida). Lycopsida med mikrofyll med sporangier i aksil.
2) Euphyllophytina som utviklet seg til planter med monopodial forgreining (hovedstengel med sidegreiner) f.eks. Psilophyton, ga bregner, frøplanter og snelleplanter (sphenopsider). Fossiler fra Euphyllophytina finnes i N-Amerika, Grønland, Kina, og Skandinavia var plassert ved tropiske strøk ved ekvator.
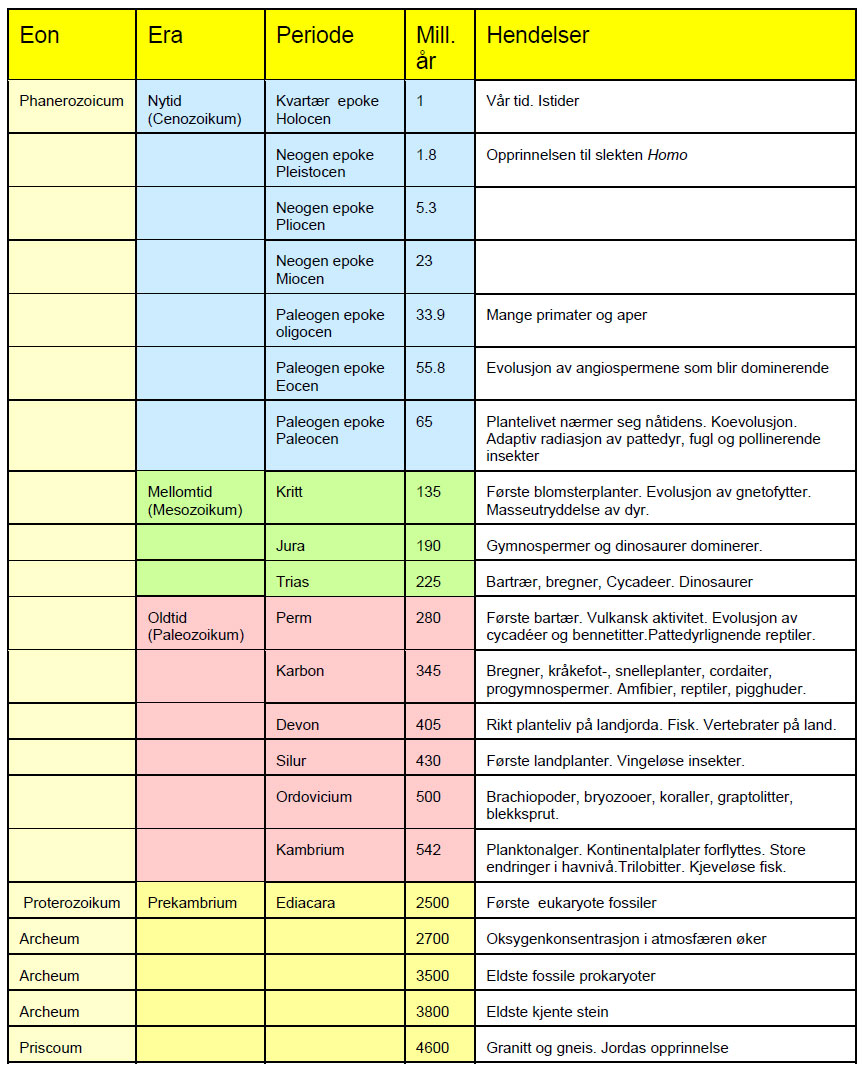
Teksten er hentet fra Evolusjon